Почки, находящиеся в пазухе пасынка, прорастают, как правило, весной, после зимовки, поэтому их называют зимующие почки, или зимующие глазки. Осенью и зимой почки винограда находятся в покое. По определению П. А. Генкеля и Е. 3. Окниной (1954), покоем у растений называется состояние, при котором почти полностью приостанавливаются процессы роста, снижается интенсивность обмена веществ и содержимое клеток претерпевает глубокие изменения.
Хотя значение пониженной температуры осенне-зимнего периода остается не вполне выясненным для дальнейшего развития зимующих почек, из работ многих авторов можно заключить, что если почки не подвергнуть воздействию пониженных температур, то они не приобретут способности к прорастанию весной и не будут развиваться дальше (Кондо, 1955). Таким образом, воздействие холодом рассматривается как средство прекращения периода покоя.
При выдерживании ранней осенью черенков винограда в холодильнике при температуре, близкой к 0°С, органический покой почек длится всего около 1,5 месяца. Растянутость периода органического покоя в естественной обстановке на виноградниках объясняется тем, что в природных условиях температура сохраняет свое оптимальное для прохождения покоя значение ограниченное время. Осенью (особенно в дневные часы) она долго держится выше необходимого оптимума, а зимой — ниже.
Я. И. Потапенко и В. А. Костина (1950) считают, что для приобретения свойства прорастания почки должны подвергнуться действию пониженной температуры в пределах 8— 0° С. Продолжительность действия пониженной температурой специфична для отдельных сортов и обусловливается их эколого-географическим происхождением. Распускание почек ускоряется тем больше, чем дольше зимой, до перемещения в теплицу, они остаются в условиях открытого грунта (Magoon, Dix, 1943). Сопоставление метеорологических условий с датой распускания почек кустов, зимовавших в открытых условиях, показало, что с увеличением холодного периода уменьшается количество тепла, необходимого для распускания почек (Branas, Bernon, Levadoux, 1946). Температура в пределах от 1 до 8°С эффективнее влияет на сокращение органического покоя, чем температура на уровне 0°С, однако температура 18 или 25° еще эффективнее (Nigond, 1957).
Воздействие температурой 5 пли —5°С в течение 15 дней не сокращает срок прорастания почек и не повышает процент распустившихся глазков (Alleweldt, 1960). К аналогичным выводам приходят также и другие авторы (Antcliff, May, 1961).
Ряд исследователей (Потапенко, 1949; Потапенко, Костина, 1950; Тютюник, 1953) считают, что пониженные температуры начинают действовать на почки очень рано — еще в конце лета и начале осени.
Необходимо отметить, что не выяснен вопрос о параметрах пониженной температуры, которая оказала бы требуемое физиологическое действие на прорастание почек весной. В работах В. 3. Гулнсашвилп (1954) указано, что продолжительность действия пониженной температуры составляет 57 дней, причем не требуется, чтобы температура падала ниже ГС.
Как выяснилось впоследствии, для прорастания почек до вступления их в состояние покоя не обязательно прохождение периода пониженной температуры. А. С. Мержаниан (1939) давно сообщал, что прищипка верхушек побегов и удаление пасынков примерно до конца июля вызывают развитие зимующих почек в год их формирования. А. М. Негруль (1952) также отмечает, что при удалении всех точек роста можно вызвать развитие зимующих почек в год их формирования.
К. Д. Стоев и М. М. Ников (1956) показали, что удаление всех точек роста сортов Памид, Димят, Болгар, Мускат белый (Тамянка), Пино черный и Юни блан с 15 июня до 15 июля вызывает массовое прорастание зимующих почек. При удалении точек роста в конце июля (в начале вызревания побегов) прорастание почек уменьшилось на 15—30%, 15 августа— на 20—30%, а при удалении точек роста в середине сентября прорастали лишь единичные почки.
В более поздних опытах М. М. Никова (1964) при удалении точек роста до 10 июля прорастало 100% почек, при более позднем удалении точек роста количество прорастающих почек уменьшалось (табл. 32).
Таблица 32
Динамика прорастания зимующих почек при удалении точек роста в период 10 июля — 25 августа (в %)

М. Ников установил также, что потеря способности к прорастанию почек различных ярусов наступает не одновременно. Как правило, низко расположенные почки раньше переставали развиваться, по сравнению с почками средних и высоких ярусов. Например, почки 5-го узла сорта Димят к 25 июля 1959 г. прорастали всего на 32%, в то время как почки 15-го узла на 94%. Аналогичный вывод сделал также И. Н. Кондо (1960).
О том, что для прорастания зимующих почек не требуется пониженной температуры, свидетельствует также то, что в условиях тропиков ритмы роста и плодоношения винограда осуществляются без действия низких температур (Cheema, 1954; Vega Jose, 1961; Шахова, 1961; Негруль, Мохова, 1964).
А. М. Негруль и Е. И. Мохова (1964) показали, что в тепличных условиях Москвы без искусственного освещения, но при постоянной высокой температуре можно закончить годовой цикл винограда примерно за 9 месяцев. Продолжая наблюдения в течение нескольких лет (1952—1957 гг.), они могли установить состояние покоя и вызвать рост почек в любое время года (рис. 24).

Рис. 24. Период вегетации и покоя у винограда в условиях теплицы:
А — амурский обоеполый (сеянец № 44), пониженная температура до осени 1955 г.; Б — амурский обоеполый (сеянец № 4), без пониженной температуры; В — Варандеин (сеянец № 58), пониженная температура до осени 1955 г.; Г — Варандеин (сеянец № 24), без пониженной температуры; в — вегетация; п — покой (числа около букв в и п показывают число дней вегетации или покоя)
Конец вегетации наступал всегда, как только заканчивался рост побегов в длину и толщину и начиналось их вызревание. Зимой, как правило, почки находились в состоянии покоя более продолжительное время, чем летом. Ритмы вегетации осуществлялись независимо от длины дня.
Потеря способности зимующих почек к прорастанию вызвана их вступлением в состояние покоя (непрорастания). Как выяснилась, в состояние покоя почки винограда на европейском континенте входят с конца августа — начала сентября до конца октября (Кондо, 1955; Nigond, 1957; Alleweldt, 1960), а «а американском (Калифорния) на месяц позже (Winkler, 1962). В южном полушарии в состояние непрорастания почки вступают в апреле—мае (Winkler, 1962; Негруль, Мохова, 1964).
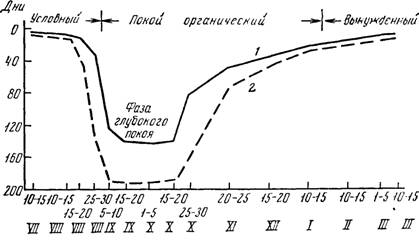
Рис. 25. Динамика состояния покоя почек винограда: / — среднеазиатские сорта; 2 — европейские сорта
И. Н. Кондо (1955) изучал явление непрорастаемости почек в условиях Узбекистана и юга Казахстана, пользуясь методом проращивания в лабораторном помещении. Вступившими в состояние органического покоя он считал почки, которые не прорастали в течение минимум 20 дней, а вышедшими из этого состояния те, которые прорастали менее чем за 20 дней. На основе многолетних исследований И. Н. Кондо выразил динамику состояния почек графиком (рис. 25).
Как видно из графика, покой почек бывает условным, органическим и вынужденным. В зависимости от местоположения зимующих почек по длине побега, района выращивания и сорта винограда, условный покой может проявляться в северном полушарии до начала августа — начала октября. В состоянии условного покоя находятся замещающие почки глазка, спящие почки и нераспустившиеся глазки однолетней лозы. Условный покой легко можно прервать и вызвать прорастание почек.
Состояние органического покоя выражено наиболее сильно в начальном этапе вступления почек в состояние непрорастания, которое выделяется как фаза глубокого покоя. Общая продолжительность органического покоя у сортов разного эколого-географического происхождения неодинаковая — у восточных 4—4,5 месяца, а западных и черноморского бассейна 4,5—5,5 месяца.
В период вынужденного покоя состояние непрорастания почек вызвано наличием неблагоприятных условий внешней среды.
И. Н. Кондо (1959) показал, что в орошаемых условиях почки у основания побега могут вступать в состояние покоя еще до того, как рост прекратится. В условиях засухи, наоборот, почки вступают в состояние непрорастания после окончания роста. Таким образом, рост побегов и вступление почек в состояние непрорастаемости не исключают один другого.
Длительность периода покоя почек сорта Султанина в условиях Австралии около 3 месяцев, т. е. значительно меньше, чем было установлено И. Н. Кондо (1955, 1959) для европейских и азиатских сортов, что, по-видимому, связано с условиями их произрастания. Осенью и зимой в Австралии также наступает непрерывное уменьшение глубины покоя. Ранняя осень — период наиболее глубокого покоя, интенсивность которого уменьшается с наступлением холодного периода. Органический покой можно ясно разграничить от вынужденного лишь в фазе глубокого покоя. При переходе органического покоя в вынужденный их трудно разграничить и выбор разделяющего их пункта произволен (Antcliff, May, 1961).
Пуже (Pouget, 1960, 1961, 1963) разделяет период, в течение которого почки находятся в состоянии потенциального роста, на следующие фазы.
1. До покоя: в этой фазе почки начинают распускаться после очень короткого периода (10—40 дней).
2. Покой: характеризуется тем, что срок прорастания почек по сравнению с фазой до покоя увеличивается на 50%, причем это свойство наступает и достигает максимальной величины сразу.
3. Прекращение состояния покоя: выход из покоя наступает резко, как и вступление в него и характеризуется тем, что срок прорастания почек снова снижается на 50%.
4. После покоя.
5. До бутонизации.
Вместе с изучением ритмических явлений развития почек винограда все большее место занимают исследования, посвященные средствам и методам подавления или ускорения выхода зимующих почек из состояния непрорастания. Из всех средств чаще всего испытывали хлоргидрин, индолилуксусную кислоту, гиббереллин, риндит и др.
Этилен хлоргидрин и индолилуксусная кислота выводят зимующие почки из состояния непрорастания, причем эффект тем более заметен, чем раньше проводят обработку; гпббереллин, наоборот, удлиняет период непрорастания (Antcliff, May, 1961) (табл. 33).
Подавление состояния прорастания почек сорта Султанина гиббереллином, по мнению Антклифа и Мея,— новое доказательство того, что точкам сорта не требуется похолодания, чтобы выйти из состояния покоя. Гиббереллин заменяет холод и выводит почки из состояния покоя таких видов растений, которым для этого необходимы пониженные температуры (Brian, 1959).
Таблица 33
Процент распустившихся почек и срок их прорастания при 20° С под влиянием обработки различными веществами (одноглазковые черенки)
Вещество, использованное для обработки почек | Процент распустившихся глазков | Срок до прорастания, дни |
Контроль | 99,2 | 27,3 |
Этилен хлоргидрин | 98,3 | 23,7 |
ИУК | 99,2 | 26,9 |
Гиббереллин | 61,7 | 85,1 |
Обработка почек а-нафтилуксусной кислотой угнетающе действует на их рост, так же как и наличие терминальной почки. В этом дополнительное подтверждение гипотезы, что взаимное подавление прошлогодних почек, развитых на одном побеге, имеет гормональный характер (Bessis, 1963).
Ранняя обработка почек (в октябре и ноябре) а-нафтил-уксусной кислотой не оказала практического влияния на прорастаемость почек (Nigond, 1960). Видимое подавление прорастаемости почек установлено при обработке почек между 15 января и 25 февраля, что приводило к задержке распускания почек на 16—27 дней (рис. 26). Для получения эффекта срок обработки имеет большее значение, чем концентрация реагента.
Торможение прорастания почек а-нафтилуксусной кислотой — перспективный метод искусственного удлинения срока распускания почек для предупреждения повреждений их поздними весенними заморозками (Nigond, 1960).
Сильно подавляющее (при определенных концентрациях и сроках обработки) действие на зимующие и летние почки оказывает малеиновый гидрацид (Huglin, Julliard, 1959).
Наиболее популярный метод стимулирования выхода зимующих почек из состояния покоя и ускорения их прорастания — обработка теплой водой. Метод этот известен и широко применяется в цветоводстве и некоторых других отраслях растениеводства. Наблюдения показали, что обработка теплой водой (30°С) свыше 16 ч прерывает покой с большой эффективностью (разница по сравнению с контролем 80 дней). Аналогичный эффект получался и при анаэробиотическом воздействии азотом в течение 16 и 24 ч. Выдерживание почек в полиэтиленовых пленках вызывало выход из покоя лишь через 96 ч. Эффективность анаэробиоза сильнее выражена в более ранний срок (Pouget, 1961).
На основании этих исследований Пуже приходит к выводу, что анаэробные условия вызывают выход из состояния покоя. При использовании риндита как средства ускорения выхода почек из состояния покоя обнаружены существенные изменения в характере дыхания; временное ослабление интенсивности кислородного дыхания, а вместе с этим появление анаэробного дыхания и усиление гликолиза (Pougel, 1960, 1962). Если коэффициент дыхания в контрольном варианте оставался на уровне 1 (от 0,89 до 1), то в варианте, где почки обрабатывали риндитом, он всегда был выше 1 и через 5 дней после обработки достигал величины 2,45. Это можно объяснить лишь усилением анаэробного дыхания и дополнительным потреблением субстратов дыхания (углеводы).
В прекращении состояния покоя почек под действием риндита и в переходе к анаэробному дыханию Пуже видит определенную связь и считает его одним из возможных путей прерывания покоя.
Ускорение выхода из состояния покоя установлено также под действием некоторых других химических веществ: СаСЬ, ZnS04, CuS04, MgS04 и др. (Pieri, 1953).
Вступление почек в состояние покоя, как и выход из него, связано с определенным обменом веществ. Исследования показали, что во время покоя почек происходит ряд существенных изменений в содержании крахмала, липоидов, дубильных веществ и жиров, причем эти превращения имеют специфический характер и очень тесно связаны с морозоустойчивостью сортов и условиями зимовки (Окнина, Кузьмин, 1955; Окнина, Кузьмин, Молчанова, 1965).

Рис. 26. Влияние а-нафтилуксусной кислоты на развитие почек в контролируемых условиях:
1 — обработка 20 ноября 1956 г.; 2 — обработка 8 января 1957 г.; 3— обработка 23 февраля 1957 г.; 4 — контроль (без обработки)
Во время покоя почек меняется также изоэлектрическая точка (ИЭТ) протоплазмы. По данным Е. К. Плакиды и Н. О. Лагутинской (1952), рН ИЭТ почек винограда, находящихся в состоянии зимнего покоя, равна 5,4—5,8, в то время как в почке, тронувшейся в рост, она сдвигается до 3,0. Они показали также, что в зависимости от возраста почки рН ее тканей сдвигается от более кислой в менее кислую зону (на 0,8-0,3 рН). По сравнению с неплодоносными почками рН ИЭТ плодоносных находится в несколько менее кислой зоне (на 0,2 рН).
И. Москов, М. Ников и Л. Бозова (1963) показали, что содержание свободных аминокислот и сахаров в почках винограда сильно меняется во время вегетации и покоя. Больше всего аминокислот в молодых почках. Постепенно с их развитием набор и содержание аминокислот уменьшается и к началу вступления почек в состояние покоя число аминокислот уменьшается. При этом динамика аминокислот неодинаковая для почек различного местоположения по длине побега: почки 5-го узла этого состояния достигают к концу августа, а почки 20-го узла — во второй половине сентября (13/IX—1/Х).
Выход из состояния покоя сопровождается новым повышением содержания и набора аминокислот (табл. 34, 35). При этом увеличение набора аминокислот отмечается для нижних почек значительно раньше (10/XI), чем для расположенных выше (12/XII).
При сопоставлении динамики аминокислот в почках со способностью последних к прорастанию выявляется хорошая согласованность их. Потеря способности к прорастанию (вступление в состояние покоя) сопровождается уменьшением набора аминокислот в почках, а восстановление прорастаемости — повышением его (табл. 36). При этом раньше выходят из состояния покоя низкорасположенные почки, а позже — находящиеся в верхних зонах побега.
Подобное явление установили и другие авторы (Кондо, 1954; Huglin, 1958).
Изменяется также содержание сахаров в почках. Во время глубокого покоя набор сахаров сильно уменьшается (сахароза и глюкоза), а выход из него сопровождается увеличением числа сахаров (глюкоза, фруктоза, сахароза и мальтоза) .
Значительные изменения претерпевают нуклеиновые кислоты. Как показали исследования А. И. Чебана (1968), они интенсивно накапливаются в зимующих почках винограда в периоды их закладки и дифференциации и весной после выхода из состояния покоя. В период покоя же накопление нуклеиновых кислот замедляется.
С переходом почек в состояние покоя содержание кислоторастворимого органического фосфора резко снижается, а после выхода из состояния покоя — увеличивается.
Таблица 34
Динамика набора свободных аминокислот в почках винограда сорта Димят (по пятибалльной системе)
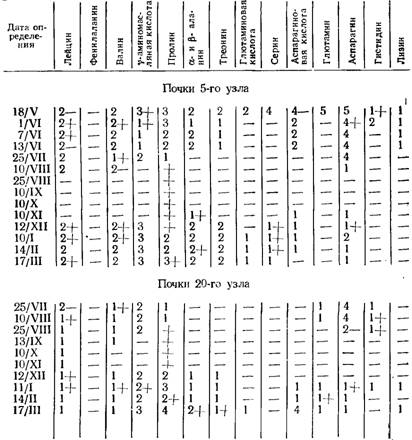
Таблица 35
Динамика набора свободных аминокислот в почках винограда

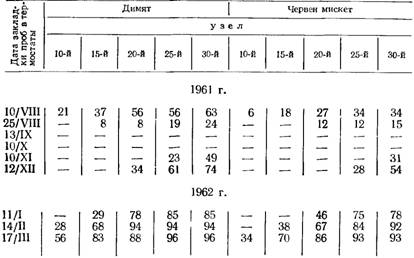
Таблица 36
Прорастание глазков у сортов Димят и Червен мискет (число проросших глазков на 10— 30-й день)
По данным В. С. Семина и В. И. Мадиса (1964), почки винограда до листопада способны поглощать неорганический фосфор и превращать его в органическую форму (фосфатидную и нуклеопротеидную).
В заключение можно сказать, что непрорастаемость (покой) почек — одно из важнейших и сложных биологических явлений, выработанное виноградом в процессе эволюционного развития как приспособительная реакция к критическим изменениям условий внешней среды. Благодаря усилиям многих ученых некоторые вопросы этого явления изучены в значительной степени. Несмотря на это, многие стороны природы покоя почек винограда до сих пор не ясны и требуют дальнейшего исследования.