Динчев и др. (1964) изучали транслокацию и метаболизм азота в листьях (4—6 и 10—12-го узла), в побегах (4-6 и 10—12-го узла) и в ягодах винограда сорта Болгар. Изучение они проводили в динамике — во время цветения, в период интенсивного роста ягод, в начале созревания ягод, во время физиологической зрелости и до наступления листопада, используя в качестве метки стабильный изотоп азота (15N).
Исследования этих авторов показали, что 15N включается во все азотные фракции листьев, причем это наблюдается на начальных этапах развития виноградной лозы. В конце вегетации (до наступления листопада) имеющийся в листьях аммиачный и нитратный азот — немеченый, следовательно, имеет почвенное происхождение.
К концу вегетационного периода (фаза физиологической зрелости винограда и до листопада) отмечается резкое уменьшение азота в запасных и, главным образом, в конституционных белках, что, по-видимому, связано с передвижением азота из листьев к побегам и к ягодам винограда. В период физиологической зрелости винограда и до листопада концентрация l5N уменьшается в пять раз по сравнению с его концентрацией в период цветения.
Наблюдения показали также, что поступление азота в различных фракциях в побеги винограда и его превращения имеют такой же характер, как в листьях. Более существенный обмен азота происходит в ягодах винограда.
Прежде всего следует отметить, что азот поступает в ягоды винограда главным образом в форме аммония и органического небелкового азота. Существенно еще то, что до начала созревания винограда поступающий в ягоды азот — немеченый, т. е. происходит из почвы. В фазу физиологической зрелости ягод и до листопада отмечается сильное обогащение фракции аммиачного азота и органического небелкового азота.
Источником этого азота, по всей вероятности, являются конституционные и запасные белки в листьях и побегах, которые подвержены интенсивным процессам ретроградации в последних фазах вегетации. Таким образом, исследования Динчева, Стоева и др. не только подтвердили вывод о том, что на начальных этапах вегетации азот накопляется в молодых листьях и побегах, но показали также, что к концу вегетации происходит ретроградация белков в этих органах, и азот переходит к ягодам, а по всей вероятности, и к другим органам винограда.
Известны и другие исследования, в которых изучались метаболизм веществ при внесении в почву азотных и фосфорных удобрений и полимикроудобрений (Вашадзе, 1972; Абесадзе с сотр., 1976), а также изменения содержания фосфора и его содержаний в разных органах винограда (Xачидзе и Мосашвили, 1971, 1976).
Значительный интерес представляют исследования Чкуасели (1966), изучавшего передвижение и транслокацию 32Р, 35S и 45Са в разных органах виноградной лозы. Он установил, что накопление 32Р в органах виноградного растения в течение суток неравномерно. Наибольшая активность накопления радиоактивного фосфора установлена днем (около 17 ч.). Максимальное накопление 32Р наблюдается в молодых органах. В фазе активного роста градиент распределения 32Р — восходящий, с хорошо выраженной ортостихной полярностью. По мере старения органов виноградной лозы градиент распределения становится нисходящим, причем явления ортостихной полярности не наблюдаются. По данным автора ионы калия способствуют накоплению 32Р в надземных органах виноградной лозы. Ионы же кальция, как правило, ослабляют этот процесс.
Радиоактивная сера (36S) накопляется преимущественно в листьях и в значительно меньшей степени в побегах. Во всасывающих корнях накопляется больше 35S, чем в проводящих. Ортостихной полярности в распределении серы Чкуасели (1966) не наблюдал.
Подобно фосфору, кальций (45Са) накопляется больше в молодых органах — в пасынках, в верхних молодых листьях и в верхушке побега, причем градиент накопления восходящий.
Хачидзе и Мосашвили (1976) изучили поступление в корни, передвижение и метаболизм 32Р в разных органах винограда при 5—7 и 27й- 28 °С. Они установили, что в зимний период на протяжении 48 h основное количество усвоенного фосфора содержалось в корнях, причем 1/3 усвоенного фосфора представлена в минеральной форме, а 2/3 — в органической. Летом при температуре 27—28 СС усвоение 32Р происходит в несколько раз активнее, чем в зимний период. Из общего количества фосфора больше всего его оказалось в листьях, затем в побегах, корнях и штамбе, а 32Р, поступая в корни виноградной лозы, активно включается в органические соединения — радиоактивность минерального фосфора ниже, чем радиоактивность органических фракций. Общая радиоактивность в старых листьях выше, а включение радиоактивного фосфора в органические соединения происходит интенсивнее в молодых листьях.
В последнее время проблема транслокации минеральных веществ в разных органах виноградной лозы изучалась весьма обстоятельно Стоевым и Бадъром (1978). Они определяли содержание N, P, K, Са, Na, Μn, Mg, Fe, Zn и Сu в листьях и побегах сорта Болгар (в нескольких секторах по длине побега), в двухлетних и многолетних ветвях и корнях диаметром до 3 mm и более. Исследование показало, что по характеру передвижения и их транслокации в разных органах винограда на протяжении годичного цикла минеральные элементы можно разделить на три группы.
В первую группу входят N, Р и Zn (рис. 6)*. Как видно, в начале вегетации эти элементы имеются в наибольшем количестве в листьях и побегах.
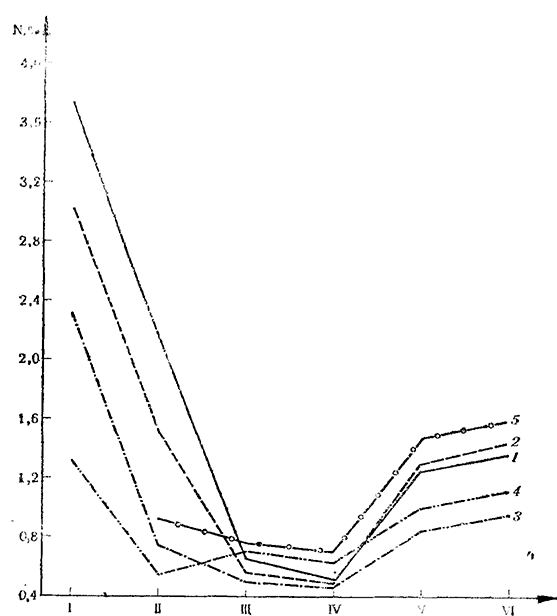
Рис. 6. Транслокация азота по фазам
1 — выделение соцветий; 2 — массовое цветение; 3 — начало созревания; 4 — начало листопада; 5 — покой; I — листья; II — побеги; III — двухлетние ветви; IV — многолетние части; V — корни <3 mm; VI—корни > 3 mm
В конце вегетации они сконцентрированы преимущественно в корневой системе (в основном, в корнях диаметром до 3 mm), а в начале нового вегетационного периода они передвигаются к молодым растущим частям куста. Двухлетние и многолетние части выявляют себя преимущественно как транспортное русло при передвижении минеральных элементов в нисходящем и восходящем направлениях. На этом основании можно считать, что азот, фосфор и цинк являются жизненно важными элементами, причем они принимают активное участие в обмене веществ в молодых растущих органах — листьях и побегах и в корневой системе винограда. Обменные процессы, в которых они участвуют, неидентичны, одинаков лишь характер их динамики и транслокации.
Ко второй группе можно причислить К, Са, Mg, Μn и Сu (рис. 7,8)**.
* Ввиду того, что динамика и транслокация цинка и фосфора имеют почти идентичный ход с азотом, отдельная диаграмма для них не приводится.
** Из-за идентичности динамики и транслокации магния, калия и меди с кальцием и марганцем их диаграмма не приводится.
Главное и общее здесь то, что их передвижение к корневой системе в осенне-зимний период очень слабо заметно, незаметно также передвижение в обратном, восходящем направлении. Это дает основание считать, что физиологическая роль упомянутых элементов в корневой системе винограда не связана с активным обменом веществ в корнях.

Рис. 7. Транслокация кальция по фазам (обозначения те же, как на рис. 6)
Наоборот, все эти пять элементов (К, Са, Mg, Μn и Сu) находятся в значительных количествах в листьях и побегах в начале вегетации и их содержание резко уменьшается одновременно с понижением активной жизнедеятельности растений. Поэтому можно принять, что К, Са, Mg, Μn и Сu активно участвуют в обмене органических веществ в листьях и побегах, несмотря на то, что обменные процессы для каждого из них специфичны. Здесь так же, как и в первой группе элементов, идентичен лишь характер их динамики и транслокации.

Рис. 8. Транслокация магния по фазам (обозначения те же, как на рис. 6)

Рис. 9. Транслокация натрия по фазам (обозначения те же, как на рис. 6)
В третью группу входят Na и Fe. В начале вегетации и в более поздние фазы развития Na и Fe накопляются интенсивно в корнях и слабее — в двухлетних и многолетних побегах (рис. 9 и 10). Из этого следует, что корневая система в период осенне-зимнего покоя выполняет функции вместилища натрия и железа. В начале следующего вегетационного периода натрий и железо передвигаются из корневой системы в восходящем направлении.

Рис. 10. Транслокация железа по фазам (обозначения те же, как на рис. 6)
Поведение обоих элементов, однако, неидентично в листьях и побегах — в начале вегетации они богаче натрием, а беднее железом. Это дает основание полагать, что натрий принимает активное участие в процессах обмена веществ в надземных органах и корневой системе, в то время как физиологические функции железа сосредоточены преимущественно в корнях.
Приведенные выше данные Дают основание считать, что N, P, K, Са, Na, Mg, Μn, Zn, Fe и Си выполняют разные физиологические функции в обмене веществ винограда. Азот, фосфор и цинк принимают активное участие в обмене веществ в листьях, побегах и корневой системе. Это создает предпосылки для интенсивного их передвижения в нисходящем направлении в конце вегетации и в восходящем — в начале роста, во время цветения и до начала созревания винограда. Эти элементы можно считать сильно подвижными. Они входят в состав важных ферментных систем и органических веществ, специфических для органов, отличающихся наиболее активным обменом — листьев, побегов и корней винограда.
Калий, кальций, магний, марганец и медь принимают участие преимущественно в процессах обмена в листьях и побегах. Их передвижение к корневой системе в конце вегетации не выражено четко, незаметно также их передвижение в обратном направлении. Эту группу минеральных элементов можно считать менее подвижной (инертной). Исследования физиологической роли К, Са, Mn, Mg и Сu показывают, что они принимают участие в регулировании ферментных систем и в обменных процессах, локализованных преимущественно в надземных органах — фотосинтезе, дыхании, транспирации, росте и передвижении ассимилятов.
Натрий и железо интенсивно передвигаются к корневой системе и менее активно — к двухлетним и многолетним стеблевым частям. Это дает основание заключить, что они участвуют в процессах обмена, протекающих преимущественно в корневой системе. Натрий принимает некоторое участие в обмене веществ также в молодых растущих органах виноградной лозы — листьях и побегах. Натрий и железо по своей подвижности занимают среднее место между первой и второй группами элементов.
В заключение следует отметить, что в динамике и транслокации азота и других минеральных веществ (Р, К, Са, Na, Mg, Mn, Fe, Zn и Сu) в органах виноградной лозы листья, молодые побеги и корни являются полюсами, определяющими направление их передвижения и локализацию. Эти процессы имеют специфический характер, ввиду чего классификация их подвижности и участия в метаболизме виноградной лозы в некоторой степени имеет условное значение. Сущность процессов обмена веществ, в которых принимают участие азот и другие минеральные элементы, должна быть объектом дальнейших исследований.