Результаты и их обсуждение
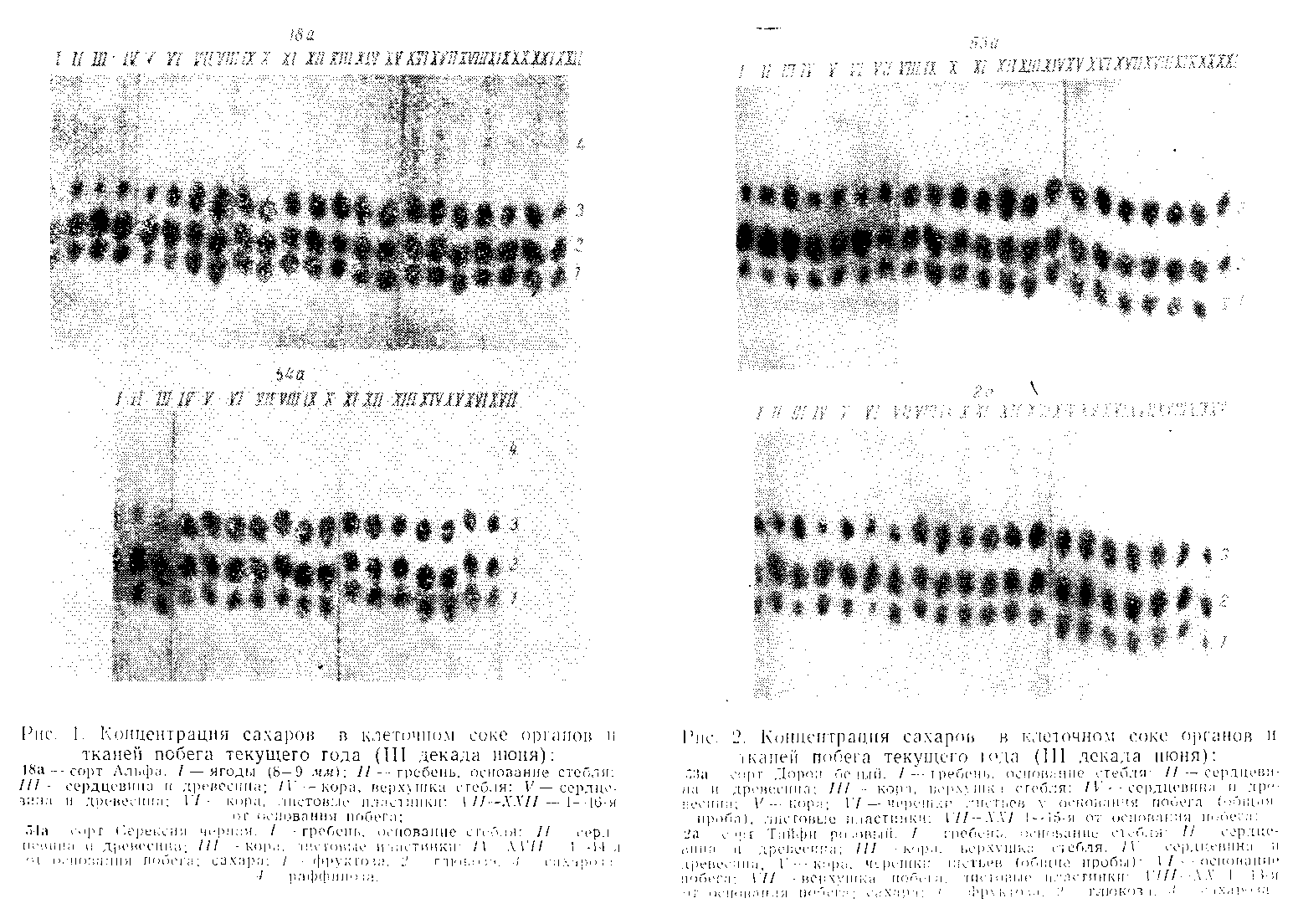
Результаты проведенного в 1965 г. изучения сахаров представлены в виде фотографий хроматограмм, позволяющих сопоставить величины концентраций анализируемых соединений в клеточном соке по органам и тканям побега, а также по срокам и сортам способом полуколичественного анализа. Данные, полученные для молодых побегов (рис. 1 и 2), показывают, что в пластинках растущих листьев по сравнению с уже сформировавшимися концентрация сахаров более низкая. С возрастом концентрация сахаров в пластинках возрастает и в дальнейшем — при пожелтении листьев — падает.
Из анализа полученных данных (рис. 1—5) следует, что первые 10—16 листьев на побеге развиваются при пониженном отношении сахарозы к моносахарам, тогда как в выше расположенных листьях сахароза превалирует над простыми сахарами. Пониженное отношение сахарозы к моносахарам, указывающее на гидролитическую направленность превращения сахаров, в самых нижних (2—4) листьях может несколько повышаться (сорт Альфа) или даже сменяться соотношением с синтетической направленностью (сорт Тайфи розовый). На основе данных по составу сахаров в пластинках листья нормального плодоносящего побега можно разделить на три яруса следующим образом: нижний — первые 2—4 листа, средний — от 3—5-го по 13—16-й лист и верхний—листья, расположенные на побеге с 14—17-го узла до его верхушки. Число листьев верхнего яруса, следовательно, достигает у закончившего рост побега 20 и более. У сортов Дорой белый и Серексия черная нижний ярус нельзя выделить столь же отчетливо по величине соотношения сахаров.

Разделение листьев побега на ярусы, исходя из соотношения сахаров в них, хорошо согласуется с данными по характеру развития листьев. Так, все наиболее крупные на побеге листья оказались включенными в средний ярус, кривые увеличения площади пластинок, т. е. их
прироста, у листьев среднего яруса имеют более крутой ход но сравнению с листьями нижнего и верхнего ярусов и т. д.
Кроме общей закономерности в изменении отношения сахароза — моносахара но ярусам листьев, весьма четко прослеживаются более или менее значительные колебания этого отношения внутри ярусов.
Особенно заметное периодическое ослабление гидролитической направленности превращения сахаров и нарастание синтетической наблюдается в нижнем и среднем ярусах. Так, у сорта Альфа в среднем ярусе почти отсутствуют периодические увеличения отношения, но у Серексии черной они резко выражены.
Сравним данные, полученные для самых нижних на побеге пяти листьев и верхушечных, взятых в виде общих проб в конце июля, в период созревания ягод у ранних сортов (рис. 3 и 4). Если верхушечных листьях обнаружено высокое отношение сахарозы к моносахарам для всех сортов, то в нижних листьях величина этого отношения позволяет говорить о различии между сортами. Раннеспелые сорта Жемчуг Саба и Дорой белый имеют низкие значения этого показателя, а поздний сорт Тайфи розовый — высокое. Поздний сорт Серексия черная, у которого ягоды созревают немного раньше, чем у Тайфи розового, занимает промежуточное положение. Следовательно, согласно этому показателю сорта различаются по степени их раннеспелости. Этот вывод подтверждается также данными, полученными еще в период цветения— завязывания ягод (см. рис. 1 и 2). Так, у раннего сорта Дорой белый отношение сахароза — моносахара имело пониженное значение для всех нижних листьев, а у позднего сорта Тайфи розовый, наоборот, наблюдалось высокое отношение в каждом листе нижнего яруса, что и сказалось на результатах анализа общих проб из пяти листьев. У Серексии черной при пониженном в целом отношении сахароза моносахара в нижних листьях довольно резко выраженная периодичность внутри ярусов привела к некоторому повышению результата анализа общей пробы.
Вероятно, при подобном анализе листьев раннеспелого сорта Альфа получили бы достаточно низкую величину отношения несмотря на то, что первые два листа показывают несколько повышенное отношение сахарозы к моносахарам.
Полученные данные по соотношению форм сахаров в листовых пластинках убеждают в том, что в нижних и средних на побеге листьях у ранних сортов в отличие от поздних наблюдается в целом более сильный сдвиг отношения сахароза - моносахара в сторону увеличения доли последних при общей более высокой концентрации суммы сахаров. Напрашивается вывод, что листья, характеризующиеся преимущественным накоплением моноз при общей высокой концентрации сахаров, являются наиболее продуктивными в процессе снабжения ягод сахарами.
Интересно, что пасынковые листья, так же как и верхушечные, имеют наиболее высокое отношение сахароза — моносахара (рис. 5).
В период от массового цветения винограда до начала созревания урожая у ранних сортов уровень концентрации суммы сахаров остается более высоким, чем у поздних, не только в листовых пластинках, но и, как правило, во всех органах и тканях побега. Таким образом, более высокому содержанию сахаров в ягоде отвечает и более высокая концентрация их в тканях вегетативных органов. После созревания урожая наблюдается дальнейшее повышение концентрации сахаров в основных листьях побега.
Известно, что в ягодах винограда по мере их роста и созревания содержание фруктозы относительно глюкозы возрастает. Из полученных данных следует, что в рассматриваемые фазы вегетации при старении любого органа или ткани побега в отношении глюкоза-фруктоза возрастает доля фруктозы как при общем повышении концентрации моносахаров (ягоды), так и при ее снижении (остальные органы побега) к концу вегетации. Увеличение доли фруктозы в отношении глюкоза-фруктоза происходит более быстро в органах и тканях побега у раннеспелых сортов.
В клеточном соке черешков общая концентрация сахаров значительно ниже этого показателя для листовых пластинок. Однако по характеру динамики качественного и количественного состава сахаров черешки можно сравнивать с расположенными на них пластинками листьев.
В коре, а при вызревании стебля — в лубе, в течение вегетации повышается концентрация сахарозы, раффинозы, стахиозы. Концентрация моносахаров в коре (лубе) после достижения максимального значения сильно снижается к осени. По предварительным данным рефрактометрических измерений, появление коркового слоя на стебле наблюдается при достижении концентрации сухого вещества в клеточном соке коры 9,5—10%. Интересно, что по темпу накопления сахарозы, раффинозы, а также уменьшению содержания моносахаров в кори (лубе) у основания стебля позднеспелый сорт Тайфи розовый опережает такой раннеспелый сорт, как Дорой белый уже в конце июля и особенно заметно в августе. Эти данные хроматографического анализа согласуются с наблюдавшимся на участке более ранним началом вырывания побега у Тайфи розового. Но при этом различие по тем же показателям между основанием побега и его верхушкой оставалось более значительным у позднеспелого сорта Тайфи розовый.
Сравнение ряда сортов в конце вегетации подтвердило определенную связь между концентрацией сахарозы, раффинозы и стахиозы в лубе и степенью морозостойкости сорта. Однако эта связь не ноет строгого характера, особенно в отношении сахарозы.
Вызревание стебля сопровождается повышением концентрации сахарозы при одновременном снижении содержания моносахаров также в клеточном соке древесины, но процесс этот выражен несколько слабее, чем в коре (лубе).
В клетках сердцевины по мерс их старения и отмирания происходит снижение концентрации всех форм сахаров. По этому признаку клетки сердцевины схожи с другим отмирающим к осени органом листовой пластинкой.
Величина концентрации сахаров и других водорастворимых соединений в клетке, безусловно, заключает в себе больший физиологический смысл, чем выражение содержания этих веществ относительно сухого веса материала. На это обстоятельство уже обращали внимание А. Уинклер и В. Вильямс [36], а позднее и И. Н. Кондо [7]. По вашему мнению, значение использования величин концентрации особенно возрастает при изучении вопросов физиолого-биохимической природы свойства раннеспелости. Это следует уже из того, что процесс созревания ягоды мы связываем с изменением концентрации сахаров и кислот в ее клеточном соке; с. другой стороны, связь между ягодой и остальными органами побега в отношении содержания водорастворимых соединений целесообразно искать прежде всего в их концентрации, принимая во внимание возможные механизмы передвижения веществ в растении.
Отмеченные нами особенности обмена сахаров в листьях но длине побега, а также в связи со свойством раннеспелости у винограда аналогичны полученным с другими культурами. Так, более низкое отношение скорости синтеза сахарозы к скорости се гидролиза в пластинках листьев среднего яруса по сравнению с остальными листьями побега или розетки было установлено на многих культурах (яблоня, персик, свекла, картофель, овес, хлопчатник и др.) методом вакуум-инфильтрации рядом исследователей [9, 12—15, 19, 22, 24]. Отмеченные нами для винограда периодические отклонения величин отношения сахароза—моносахара от общего уровня внутри ярусов листьев наблюдались В. И. Ниловым с сотрудниками [13, 14] у исследованных ими плодовых и травянистых растений в виде колебаний отношения синтезирующего к гидролизующему сахарозу действию Ферментов. Авторы указывают на отсутствие связи этого явления с освещенностью листьев и объясняют его изменениями направленности обмена в материнских тканях побега в моменты заложения листьев. В. И. Ниловым с сотрудниками [12] также отмечалось, что у многолетних плодовых культур в отличие от травянистых однолетних часто не наблюдается повышения величины отношения синтез—гидролиз сахарозы в самых нижних листьях.
Исследованиями Б. А. Рубина и Н. М. Сисакяна [21, 22], В. И. Нилова с сотрудниками [14] было показано, что листьям раннеспелых сортов плодовых культур свойственно более низкое отношение синтез — гидролиз сахарозы по сравнению с позднеспелыми сортами.
Из аналогичных данных для растений различных систематических групп следует [19], что отмеченные особенности обмена сахаров в связи с ярусностью листьев или раннеспелостью сорта носят общий характер и обусловлены общими закономерностями роста и развития растения.
Согласно взглядам Б. А. Рубина, И. В. Гулидовой, В. Б. Соколовой [5, 18—20. 25], из листьев с повышенной интенсивностью процессов распада сахарозы происходит более энергичный отток ассимилятов, приводящий в итоге к повышению уровня продуктивности. Это положение, как показывают результаты нашей работы, очевидно, монет быть применимо также к виноградному растению.
Уже отмечалось, что исследования особенностей обмена углеводов в связи со свойством раннеспелости у растений, как правило, выполнялись на основе применения метода вакуум—инфильтрации, который нашел возможность выяснить направленность ферментативного превращения сахаров в живых неразрушенных тканях листа. Однако еще А. Л. Керсановым [8], создавшим этот метод, а позже и другими автолами [1] было показано, что соотношение величины синтетического и гидролитического действия ферментов, найденное методом вакуум-инфильтрации в нормальных условиях жизнедеятельности, соответствует соотношению сахарозы и моносахаров в органе до опыта. Это позволяет в ряде случаев решать поставленные задачи на основе результатов непосредственного анализа химического состава тканей, а кроме того, сопоставлять эти данные с результатами отмеченных работ.
Показательны в этом отношении результаты опыта с прививкой листьев сахарной и кормовой свеклы на один и тот же подвой — корни сахарной свеклы.
В привитых листьях сахарной свеклы было найдено 10,4% восстанавливают их сахаров и 1,4% сахарозы, тогда как в листьях нее сахаристой кормовой — 6,5 и 3,0% соответственно, т. е. листья сахарной свеклы характеризовались более высоким содержанием сахаров при более низком отношении сахароза — моносахара [16]. Этого следовало ожидать, принимая во внимание выводы уже упоминавшихся работ о том, что листья высокосахаристых сортов в отличие от малосахаристых характеризуются низким отношением синтетического действия к гидролитическому.
Вместе с тем данные непосредственного анализа химического состава позволяют не только сравнивать органы и ткани побега по концентрации сахаров, но и через соотношение форм сахаров иметь довольно верное представление о направленности ферментативного превращения сахаров как в листьях, так и в других органах и тканях побега.
Известно, что ферментативные опыты in vitro часто искажают картину, наблюдающуюся в неразрушенных тканях, однако, позволяют судить о величине инвертазной активности. Проведенные нами опыты показали, что по степени уменьшения активности инвертазы органы и ткани побега расположились в следующем порядке: созревающие ягоды, зеленые ягоды, листья, гребни, черешки, древесина, кора, луб. Таким образом, наиболее высокая инвертазная активность обнаружена у ягод с минимальным содержанием сахарозы, и наиболее низкая в лубе, характеризующемся самым высоким содержанием этого сахара. Дальнейшие исследования показали, что инвертаза ягод связана клеточными структурными компонентами, т. е. является нерастворимой, и по механизму действия представляет собой трансфруктозилазу. Фермент в присутствии высокой концентрации сахарозы переносит на нее остаток фруктозы с образованием кестозы. Кестоза, так же как и сахароза, гидролизуется трансфруктозилазой при низком содержании в среде сахарозы. Следы сахара, которые можно различить на хроматограммах между раффинозой и сахарозой в пробах клеточного сока ряда органов, очевидно, принадлежат кестозе. Возможно также, что кестозой является тот неидентифицированный трисахарид, о котором говорится в литературе но физиологии винограда [11]. Rf кестозы в обычных растворителях очень близок или совпадает с таковым для лактозы.
Инвертаза, безусловно, играет важную роль в процессе созревания ягод винограда и заслуживает пристального изучения. Как показали наши опыты, в ягодах, имеющих сочную, «тающую» мякоть (сорта Мускат белый, Жемчуг Саба, Катта курган, Серексия черная), инвертазная активность более высокая, чем в ягодах с несочной, волокнистой мякотью (сорта Дорой белый, Чиляки красный, Халили дели, Изабелла).
Очевидно, в данных опытах инвертаза выступала в качестве «индикатора» степени гидролитической активности всего комплекса ферментов.
Инвертаза продолжает вызывать живой интерес исследователей и в последние годы. Следует упомянуть работы М. Хеча, К. Глазиу, И. Захера [29, 30, 34], выполненные с сахарным тростником. Авторы нашли строгую прямую зависимость между энергией роста тканей и активностью в них «кислой» инвертазы, имеющей оптимум действия в кислой области pH. Столь же строгая зависимость найдена между процессом накопления сахарозы в тростнике при его вызревании и активностью «нейтральной» инвертазы с оптимумом действия в нейтральной области pH. Опыты проводились in vitro. Отводя большую роль инвертазе в процессе накопления сахара в тростнике, авторы вместе с тем считают, что сахарозо-УДФ-трансглюкозилаза из-за термодинамических условий накопления сахара против градиента концентрации не может нести этой функции.
Касаясь механизма созревания ягод у винограда, нельзя не остановиться на основных выводах, сделанных в отношении другой сахаронакапливающей культуры — сахарной свеклы. Методом прививок удалось показать, что основная роль в накоплении сахара в корне свеклы остается за этим органом, однако реализация природных потенциальных способностей тканей корня зависит в значительной мере от величины поверхности листьев, от особенностей их развития и физиологии [16].
Метод перепрививок соцветий и других органов, безусловно, также помог бы решению ряда вопросов сахаронакопления и биохимизма раннеспелости у винограда.