ГЛАВА VII
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ПЛОДОВЫХ КУЛЬТУР, РАЗЛИЧАЮЩИХСЯ по СПОСОБНОСТИ РЕГЕНЕРАЦИИ КОРНЕЙ
1. ОБРАЗОВАНИЕ И ИЗМЕНЕНИЕ В СОДЕРЖАНИИ АУКСИНОВ Й" ИНГИБИТОРОВ В ПОБЕГАХ ПЛОДОВЫХ КУЛЬТУР В ГОДИЧНОМ ЦИКЛЕ РАЗВИТИЯ
В настоящее время литература располагает достаточными сведениями относительно различий морфолого-анатомического и физиолого-биохимического характера у черенков трудно- и легко- укореняемых культурных растений, в связи с обработкой их стимуляторами роста [Максимов, 1946; Демитрадзе, 1951; Александров, Савченко, Демитрадзе, 1951; Зёдинг, 1955; Турецкая, 1961; Sew, Bose, 1966; N,anda, Purohlt, Adarsch, 1967; Kaminski, 1967].
Исследованиями Турецкой (1961), Дубровицкой (1961), Комиссарова (1962), Тарасенко (1967) установлено, что образование корней зависит от возрастного и физиологического состояния материнского растения и самих черенков. В комплекс факторов, определяющих физиологическое состояние черенков входит и содержание природных регуляторов роста, ауксинов и ингибиторов, которое способствует регенерации корней. Установлено, что немаловажной причиной неукореняемости некоторых плодовых культур является постоянство состава эндогенных ингибиторов роста и отсутствие у них реакции на обработку синтетическими ростовыми препаратами [Турецкая, Кефели, 1966; Чайлахян, Саркисова, 1968, 1970, 1971].
К такому заключению нас подвели результаты исследований, проведенных в весенний и ранне-осенний периоды развития плодовых культур. В последующих наших исследованиях изучалась динамика образования и накопления эндогенных, свободных регуляторов роста — ауксинов и ингибиторов в годичном цикле развития растений с целью установления того, является ли превалирование эндогенных ингибиторов роста над ауксинами у трудно- укореняющихся культур величиной постоянной и нет ли периодов развития растений, когда это соотношение меняется в сторону превалирования содержания стимуляторов.
Весьма справедливым является высказывание Кренке (1950) о том, что: «Для трудноукореняющихся черенками (например, яблоня) или редко образующие каллусовые придаточные побеги (например, картофель, хлопчатник и др.) важнее найти определенную фазу развития, на которой растения оказываются более способными к регенерации, чем воздействовать тем или иным стимулятором на поверхность ранения или небольшую часть побега».
На закономерные изменения содержания стимуляторов роста и ингибиторов в течение вегетации указывают некоторые исследователи [Зёдинг, 1965; Кузина, 1970 и др.]. Зёдингом в течение зимнего покоя в почках обнаружены неактивные ростовые вещества, играющие роль запасных ростовых веществ. Генке и Шмут (1965) в покоящихся почках яблони обнаружили большое количество ингибиторов роста, которое уменьшалось с наступлением вегетации, в марте — апреле.
Наши исследования проводились с теми же плодовыми культурами и сортами плодовых пород, что и в предыдущих опытах. Взятие растительных образцов производилось ежемесячно, в течение круглого года с одного и того же дерева.
Результаты этих исследований дают нам возможность судить об уровне содержания эндогенных регуляторов роста (ауксинов и ингибиторов) и их изменения в связи с прохождением различных фаз развития в годичном цикле. Как видно из рис. 39, 40, на которых представлены результаты хроматографического анализа биологических проб черенков груши и яблони, относящихся к первой группе неукореняющихся пород, до наступления вегетации (март), в экстрактах содержатся ингибиторы роста с 40% степенью торможения. Эти ингибиторы у обеих культур расположены в зонах Rf = 0,1 — 0,55 и Rf = 0,6. С наступлением весеннего сокодвижения (апрель) в побегах яблони и груши, наряду с ингибиторами роста, уже со значительно меньшей ингибирующей способностью (5 — 20%) в зонах Rf = 0,01 — 0,4 и 0,5 — 0,9 обнаруживаются стимулирующие рост вещества. Стимулирующая их способность колеблется в пределах 15 — 30%. С наступлением цветения яблони (май) в коре побегов ауксины обнаружить уже не удается; в зонах Rf = 0,3 до 0,6 обнаруживаются ингибиторы роста, процент ингибирования которых доходит до 25. Однако у груши ингибиторы в мае в Rf = 0,5 имеют меньшую ингибирующую способность (12%) и, наряду с ними, в Rf = 0,01 — 0,4 обнаруживаются стимулирующие рост вещества. В этой же зоне ауксины появляются у яблони в июне, когда у груши они уже отсутствуют.
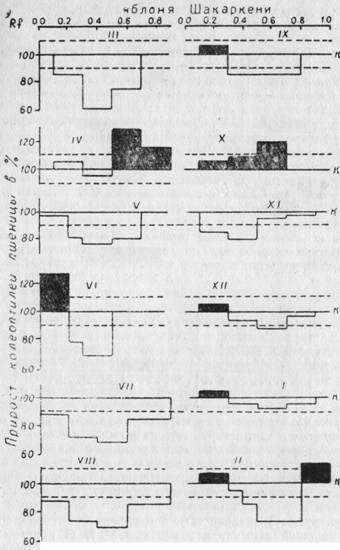
Рис. 39. Эндогенные регуляторы роста — ауксины и ингибиторы в коре и почках яблони сорта Шакаркени в годичном цикле развития. На абсциссе расположены различные величины Rf, на ординате рост колеоптилей пшеницы в % % к контролю. Пунктиром ограничен доверительный интервал (±10%).
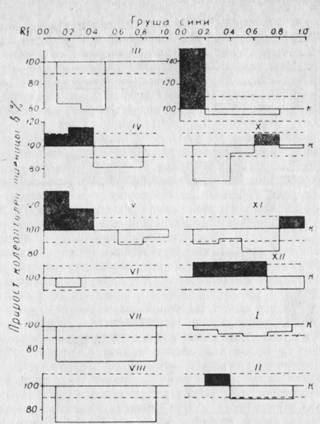
Рис. 40. Эндогенные регуляторы роста — ауксины и ингибиторы в коре и почках груши сорта Сини в годичном цикле развития. На абсциссе расположены различные величины Rf, на ординате — рост колеоптилей пшеницы в % % к контролю. Пунктиром ограничен доверительный интервал (±10%).
Высокое содержание ауксинов в коре и почках побегов текущего года обнаруживаются у груши в сентябре, в Rf=0,01 — 0,1 с 50% стимулирующей активностью, а у яблони в октябре, со значительно меньшей стимуляцией роста (20%)- С наступлением осенне-зимнего покоя следы ауксинов в различных зонах хроматограммы встречаются до февраля месяца с очень низкой стимуляцией роста (до 10%). Однако наличие следов ауксинов в осенне-зимний период сопровождается большим содержанием ингибиторов роста.
Надо отметить, что хотя и эти две культуры несколько различаются по времени образования ауксинов и их стимуляционной способности, они обладают и весьма важными общими свойствами. Эти свойства заключаются в том, что у обеих культур в побегах в течение всего годичного цикла обнаруживаются свободные ингибиторы роста. Ауксины в них встречаются в период распускания почек и цветения. С наступлением фазы плодообразования ауксины в побегах исчезают и, вероятно, транспортируют к наиболее растущим органам — плодам и точкам роста. Так продолжается до наступления осеннего периода. После съема плодов (сентябрь — октябрь) в побегах обнаруживается значительное количество ауксинов при почти полном отсутствии свободных ингибиторов роста. Такое явление продолжается недолго и уже в октябре и ноябре происходит накопление ингибиторов роста, которые не исчезают до наступления следующей вегетации. Общее явление для этих двух культур заключается в том, что соотношение ауксинов и ингибиторов в годичном цикле развития большую часть времени превалирует в сторону более высокого содержания ингибиторов роста, за исключением апреля и октября у яблони, а также мая и сентября у груши.
Хроматографический анализ и биологические пробы экстрактов из побегов плодовых культур, относящихся ко второй группе (персик и абрикос) показали, что эндогенные ингибиторы роста в них встречаются в течение круглого года (рис. 41, 42). Однако степень ингибирования этих веществ сильно изменяется в зависимости от фазы развития растения. Наивысшая степень ингибирования у этих культур отмечается в феврале и марте. В этот период ауксины в экстрактах почти отсутствуют. Относительно высокое содержание ингибиторов роста отмечается и в июне (25 — 35%). Но такое содержание ингибиторов роста сопровождается и высоким содержанием ауксинов в зонах с Rf = 0,05 — 0,3 (26 — 36%). Характерным для культур, входящих в эту группу, является то, что в сентябре в экстрактах из коры и почек обнаружены ауксины с очень высокой степенью стимулирования роста (68 — - 89%), расположенные в Rf ==0,1 — 0,3 и следы ингибиторов роста. Хотя и состав этих стимуляторов роста нуждается в дальнейшей идентификации, их можно отнести по Rf пятна и цветным реакциям к индольным соединениям. В побегах персика невысокое содержание стимуляторов роста (25%) обнаруживается в ноябре и декабре, с января по апрель обнаружить их уже не удается. В побегах абрикоса, после наступления фазы покоя, ауксины исчезают и вновь появляются лишь в период наступления вегетации.
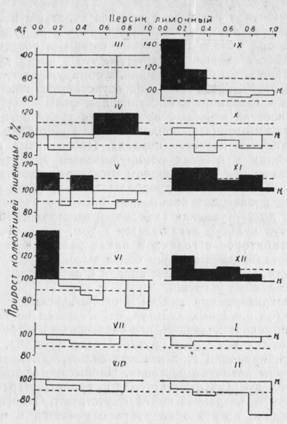
Рис. 41. Эндогенные регуляторы роста — ауксины и ингибиторы в коре и почках персика сорта Лимони в годичном цикле развития. На абсциссе расположены различные величины Rf, на ординате — рост колеоптилей пшеницы в % % к контролю. Пунктиром ограничен доверительный интервал (±10%).

Рис. 42. Эндогенные регуляторы роста — ауксины и ингибиторы в коре и почках абрикоса сорта Еревани в годичном цикле развития. На абсциссе расположены различные величины Rf, на ординате — рост колеоптилей пшеницы в % % к контролю. Пунктиром ограничен доверительный интервал (±10%).
Как видно из изложенного и приведенных гистограмм, соотношение ауксинов и ингибиторов у персика и абрикоса несколько выше, чем у представителей первой группы неукореняющихся пород. Кроме того, высокий процент стимулирования роста эндогенными ауксинами в сентябре дает возможность предположить, что в этот период можно с большим успехом добиться укоренения черенков указанных культур, чем в другое время. Такое предположение в некоторой мере обосновано и практическими результатами наших исследований.
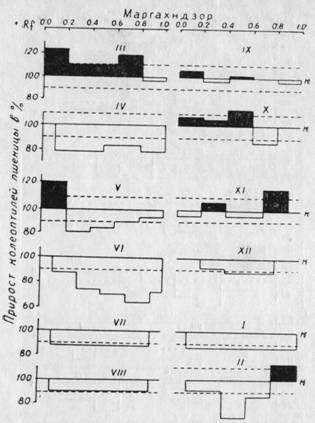
Рнс. 43. Эндогенные регуляторы роста — ауксины и ингибиторы в коре и почках карликовой яблони Маргахндзор в годичном цикле развития. На абсциссе расположены различные величины Rf, на ординате — рост колеоптилей пшеницы в %% к контролю. Пунктиром ограничен доверительный интервал (±10%).
В черенках плодовых культур, входящих в третью группу, в экстрактах в течение годичного цикла также обнаруживаются ингибиторы роста (рис. 43, 44). Ауксины, по-видимому индольной природы, в экстрактах из черенков айвы обнаруживаются в зонах Rf = 0 1 — 0,4 в мае, июне и сентябре. При этом, в сентябре степень стимулирования достигает 94 %. В этот период, как показали наши опыты предыдущих лет, и было получено 36% укоренения черенков айвы.
Несмотря на то, что по способности укоренения черенков карликовая яблоня не отличается от айвы, в составе эндогенных регуляторов роста у них есть существенные различия: у карликовой яблони обнаружены фенольные соединения, отсутствующие в тканях нормальной яблони.

Рис. 44. Эндогенные регуляторы роста — ауксины и ингибиторы в коре и почках айвы местной в годичном цикле развития. На абсциссе расположены различные величины Rf, на ординате — рост колеоптилей пшеницы в % % к контролю. Пунктиром ограничен доверительный интервал (±10%).
Наивысшая степень ингибирования роста у карликовой яблони отмечается в июне и в феврале. Ингибиторы эти располагаются по всей хроматограмме (Rf = ot 0,1 до 0,95). В феврале кроме ингибиторов обнаруживаются и рост стимулирующие вещества, которые, по-видимому, относятся к простым фенолам. В отличие от всех других культур, в экстрактах из побегов карликовой яблони ауксины обнаруживаются еще до наступления сокодвижения (март) в Rf = 0,1 — 0,8. Затем они появляются в мае, октябре и ноябре. Как видно из рис. 43, содержание ингибиторов имеет связь не только со способностью черенков к образованию корней, но и к другим процессам, определяющим карликовость этого вида яблони, т. е. общую подавленность ростовых процессов.
Проведенные нами исследования и результаты, изложенные в данном разделе, говорят о том, что корнеобразование у черенков — очень сложный процесс и зависит как от совокупности внешних, так и от внутренних физиологических факторов.
Из всех физиологических процессов, которые были изучены нами с целью выявления причин трудной укореняемости черенков плодовых культур, произрастающих в условиях сухого и жаркого климата Армении, содержание эндогенных ингибиторов и ауксинов играет первостепенную роль. По содержанию эндогенных регуляторов роста плодовые культуры резко различаются между собой. Это различие отражается и на способности их черенков к укоренению.
У культур, относящихся к неукореняющимся, обнаружено в годичном цикле развития количественное превалирование эндогенных ингибиторов роста над ауксинами, что препятствует стимулированию корнеобразования их черенков.
Эндогенные ингибиторы роста встречаются и в побегах плодовых культур, входящих во вторую группу, однако степень ингибирования этих веществ очень сильно меняется в зависимости от фазы развития растений.
У культур, относящихся к третьей группе, также наблюдается накопление в побегах эндогенных ауксинов, стимулирующий эффект которых достигает до 94%.
Таким образом, очевидно, что уровень содержания эндогенных физиологически активных веществ, ауксинов и ингибиторов, вырабатываемых самими растениями, и их взаимодействие с синтетическими ростовыми препаратами, вводимыми извне, зависят не только от физиологического и биологического состояния растения, но и от количественного их соотношения [Чайлахян, Саркисова, 1968, 1970]. К такому заключению приходит и Шпигель [Spiegel, 1954], который считает, что одной из причин трудности образования корней у черенков некоторых сортов винограда является наличие большого количества ингибиторов. Легкоукореняющиеся сорта винограда и их гибриды в исследованиях Шпигеля содержали небольшое количество ингибиторов и больше — стимуляторов роста. Такое соотношение ингибиторов над ауксинами установлено нами у ряда трудноукореняющихся плодовых культур.