В.А.Волынкин, д.с.-х.н. начальник отдела,
В.А.Зленко, н.с., к.с.- х.н.,
A. А.Полулях, с.н.с., к.с.-х.н.,
Н.П.Олейннков, в.н.с., к.с.-х.н.,
B. В.Лиховской, зав. сектором питомниководства отдел селекции, генетики винограда и ампелографии Национальный институт винограда и вина «Магарач»
РЕЗУЛЬТАТЫ ЭКСПЕРИМЕНТАЛЬНЫХ ИССЛЕДОВАНИЙ ФОРМИРОВАНИЯ ГЕНЕТИЧЕСКОГО РАЗНООБРАЗИЯ У СЕМЕЙСТВА ВИНОГРАДА VITACEAE В ПРОЦЕССЕ ЕСТЕСТВЕННОЙ ЭВОЛЮЦИИ
Проведены исследования по межродовой гибридизации (Vitis х Ampelopsis и Vitis х Parthenocissus) с применением метода скрещивания различных родов на тетраплоидном уровне. Для получения тетраплоидных сеянцев (диплоидных гамет) начавшие распускаться почки, а также соцветия за 5-14 дней до начала мейоза обрабатывали колхицином в концентрациях 0,5; 1 и 2%. С целью преодоления межродовой несовместимости, сеянцы выращивали из зародышей в культуре in vitro, выделенных из незрелых семян (40 дней после опыления). Некоторые гибридные сеянцы, выращенные в теплице, имели промежуточную степень рассеченности листьев, свойственную видам родов Vitis и Ampelopsis, которые использовались в скрещиваниях методом аллотетраплоидии.
Развитие биологических наук и биотехнологий в XX и XXI вв. позволяет создавать формы растений, которые не удавалось вывести ранее, используя только традиционный метод генеративной гибридизации. Получение таких «нетрадиционных» форм растений возможно либо на основе методов генной инженерии, либо на основе использования биологического потенциала самих растений, который заложен в процессе естественной эволюции. Эволюция у представителей родов семейства Vitaceae происходила в различных эколого-географических условиях, на разных континентах, под воздействием определенных биотических и абиотических факторов отбора и возможности естественной скрещиваемости. Если виды подрода Euvitis скрещиваются между собой легко, то получить гибриды между видом V. vinifera (подрод Euvitis, 38 хромосом) с видом Vrotundifolia Michaux (подрод Muscadinia, 40 хромосом) удалось с большим трудом [12]. Эти гибридные сеянцы стали фертильными только после их полиплоидизации (аллотетраплоидии) [13].
Ранее (1922-1924 гг.) Г.Д. Карпеченко было экспериментально доказана возможность получения фертильных межродовых гибридов редьки и капусты методом аллополиплоидии (аллотетраплоидии) [5, 6]. Но даже у растений, роды которых представлены широким спектром полиплоидных рядов, применение методов аллополиплоидии и культуры in vitro, изолированных из семян зародышей, для получения межродовых гибридов связано с большими трудностями из-за несовместимости различных родов, принадлежащих к одному семейству растений, на генетическом (генетически нестабильных) и физиологобиохимическом уровнях [7, 9]. Также генетически нестабильными являются межродовые соматические гибриды, полученные в результате слияния протопластов в культуре in vitro [3].
Искусственная межродовая гибридизация у семейства Vitaceae до наших исследований в мире не проводилась. Создание межродовых гибридов у винограда (хотя бы с частичным присутствием генов различных родов из-за возможной элиминации хромосом одного из них) позволит получить качественно новые, иммунные к биотическим и устойчивые к абиотическим факторам среды генотипы винограда, урожай которых может иметь новое направление использования, например, в фармакологической и парфюмерной промышленности.
Цель исследований - получение методом аллотетраплоидии межродовых гибридов в пределах семейства Vitaceae: скрестить род Vitis (V. vinifera) с родами Ampelopsis (A. acontifolia, A. cordata) и Parthenocissus (Р. inserta и Р. guinguefolia). Специфичность эксперимента заключалась в создании впервые в мире гибридов между сортами V.vinifera L. (род Vitis, 2n=38 хромосом) и видами других родов (Ampelopsis и Parthenocissus, 2n=40 хромосом) семейства Vitaceae: Ampelopsis acontifolia Lavalee, Ampelopsis cordata Michaux, Ampelopsis serjaniefolia Regel, Parthenocissus inserta Fritch и Parthenocissus guinguefolia Planch. [1].
При проведении исследований ставились следующие задачи:
- обработать колхицином в трех концентрациях (0,5, 1 и 2%) соцветия винограда родов Vitis, Ampelopsis, Parthenocissus за 5-14 дней до начала мейоза в клетках генеративных органов с целью образования полиплоидных гамет;
- провести инцухт у обработанных колхицином представителей различных родов и выделить семена из ягод с целью получения полиплоидных сеянцев, и после вступления их в пору плодоношения провести скрещивания между ними;
- после обработки колхицином соцветий скрестить род Vitis (вид V.vinifera L.) с видами родов Ampelopsis и Parthenocissus, выделить из зеленых ягод незрелые семена (25, 30, 40 и 50 дней после опыления) и высадить в культуру in vitro недоразвитые зародыши семян для преодоления физиологобиохимической несовместимости между родами на стадиях развития зародышей и растений из них в культуре in vitro;
- высадить на адаптацию к условиям in vivo в гидропонную культуру в теплицу растения-сеянцы, развившиеся из зародышей в культуре in vitro, которые выделены из незрелых семян (40 дней после межродовой гибридизации) и к концу вегетации вырастить из них мощные растения для пересадки в полевые условия;
- высеять в гидропонную культуру в теплице семена (полученные от различных вариантов: обработка колхицином начавших развиваться побегов, инцухт, контроль - свободное опыление) различных родов для получения полиплоидных форм, а также высев семян, полученных в результате межродовой гибридизации;
- высадить на постоянное место в поле сеянцы, полученные в вариантах после обработки колхицином, отобранные по анатомо-морфологическим признакам, а также сеянцы, полученные в результате межродовой гибридизации и развившиеся из зародышей в культуре in vitro.
Применялся традиционный путь преодоления генетической несовместимости при межродовой гибридизации (разного количества диплоидного набора хромосом у исходных форм), заключающийся в методе аллополиплоидии - использовании для скрещивания полиплоидных родительских форм. Наличие парных хромосом каждого рода у гибридных сеянцев необходимо для конъюгации между хромосомами в процессе прохождения мейоза и образования гамет, что может обеспечить фертильность гибридных сеянцев [8].
С целью получения межродовых гибридов методом аллополиплоидии (обработка растворами колхицина начавших распускаться вызревших почек или соцветий за 5-14 дней до начала мейоза для образования полиплоидных гамет) исследования проводили по двум направлениям:
- инцухт у представителей различных родов для получения из семян сеянцев - тетраплоидов и затем, после вступления в пору плодоношения, провести межродовые скрещивания тетраплоидных сеянцев;
- для ускорения межродовой гибридизации: после обработки колхицином возможно образование полиплоидных гамет (яйцеклетки и пыльцы). Поэтому проводились скрещивания между обработанными колхицином родительскими генотипами - представителями различных родов.
Так как в семенах, полученных в результате межродовой гибридизации, наблюдается гибель зародышей из-за физиолого-биохимической несовместимости различных родов, были собраны незрелые ягоды (30, 40 и 50 дней после гибридизации), которые хранились в холодильнике в течение 8-12 недель при — 2°С. Затем из ягод были выделены семена, простерелизованы 10% «Доместас» в течение 12-15 мин., затем 96%-ным спиртом 10-20 сек. И промыты 4-5 раз стерильной водой. Семена в чашках Петри в ламинарном боксе были разрезаны поперек и носики семян (части семян, в которых находятся зародыши) были высажены в три варианта жидкой среды, различающихся между собой содержанием регуляторов роста:
- 0,2 мг/л БАП для развития сердцевидных зародышей из глобулярных;
- 0,1 мг/л β-индолилуксусной кислоты (ИУК) и 30 мг/л гумата Na для превращения сердцевидных зародышей в торпедовидные;
- 0,2 мг/л гибберелловый кислоты (ГА3) для развития проростков с зелеными семядолями и гипокатилями из торпедовидных зародышей. Основа жидкой среды состояла из среды для размножения растений [4] следующего состава, макроэлементов: 308 мг/л NH4NO 922 мг/л KNO3, 597 мг/л MgSO4 7Н2О, 82 мг/л КН2РО4, 331 мг/л СаС12; микроэлементы и Fe-EDTA [11], 20 мг/л мезо-инозита, 0,5 мг/л никотиновой кислоты и 10 г/л сахарозы, но с повышенным содержанием витаминов тиамина и пиридоксина (по 5 мг/л). pH каждого варианта среды перед автоклавированием (1 атм, 25 мин) было доведено до 5,6 [2].
Проростки пересажены на питательную твердую среду для развития у них побегов и корней. Состав концентраций компонентов этой среды отличается от приведенной выше основы среды более низкой концентрацией витаминов (0,1 мг/л тиамина и 0,2 мг/л пиридоксина), добавкой 30 мг/л гумата Na, 7,5 г/л агара и концентрацией регуляторов роста: 0,15 мг/л ИУК, 0,005 мг/л а - нафтилуксусной кислоты (НУК) и 0,001 мг/л БАП; значение pH было доведено NaOH до 6,0-6,2 перед добавкой агара и автоклавированием.
Отбор полиплоидных генотипов по анатомо-морфологическим признакам у сеянцев, полученных в результате инцухта, после обработки начавших распускаться вызревших почек или соцветий растворами колхицина до начала мейоза, проводился по методике Топалэ [10].
Результаты исследований.
Получить межродовые гибриды у растений можно методом аллотетраплоидии: скрещивание исходных генотипов - представителей различных родов. С целью образования тетраплоидных сеянцев у представителей различных родов, предварительно, за 17-25 дней перед цветением (5-14 дней до начала мейоза), соцветия были обработаны (15-16 мая 2009 г.) колхицином в трех концентрациях: 0,5; 1 и 2%, с добавкой биологически активных веществ (основа растворов колхицина): 0,5 мг/л 6-бензиламинопурина (БАП), 40 г/л Д-манита, 2% димексида, 0,5 мл/л Твина 20 и 200 мг/л Na-бензоата; pH = 5,4.
Обработка соцветий этими тремя вариантами концентраций колхицина (0,5; 1 и 2 %) была проведена у родов: Ampelopsis (виды Ampelopsis acontifolia, Ampelopsis cordata), Parthenocissus (виды Parthenocissus inserta и Parthenocissus guinguefolia) и Vitis (V. vinifera миксоплоидные обоеполые сорта Харти про Ливье, Пикпуль черный, Шабаш крупноягодный и Яхеи с женским типом цветка). Затем у этих представителей различных родов был проведен инцухт (всего 24 соцветия).
В 2009 г. исследования вели в двух направлениях: инцухт у обработанных колхицином соцветий генотипов различных родов для получения полиплоидных сеянцев и после вступления их в пору плодоношения - проведение межродовых скрещиваний. В результате инцухта после обработки соцветий колхицином у вышеперечисленных генотипов получены семена (контроль - свободное опыление), всего 10800 шт. семян. Семена рассортированы по размерам, так как, по данным Ш.Г.Топалэ [10], полиплоидные семена по размерам больше, чем диплоидные, и в 2010 г. высеяны в теплицу для отбора полиплоидных сеянцев по анатомо-морфологическим признакам по методике Ш.Г.Топалэ [10]. После вступления в пору цветения между выделенными по морфологическим признакам и методом цитогенетического анализа тетраплоидными сеянцами различных родов будет проводиться межродовая гибридизация.
Второе направление исследований: проведение скрещиваний между генотипами - представителями различных родов, у которых предварительно обработаны колхицином начавшие распускаться вызревшие почки или соцветия перед началом мейоза для образования диплоидных гамет (яйцеклеток и пыльцы) и затем гибридных сеянцев - аллотетраплоидов.
При разрезании семян перед высадкой на три варианта жидких сред были выявлены их специфические сорто-биологические особенности. У сорта Яхеи при межродовой гибридизации образуются пустые семена (без зародышей и эндосперма); у сортов Шасла Гро Куляр белая и Харти про Ливье - как пустые семена, так и с эндоспермом, но проростки не развивались, а у сорта Пикпуль черный почти все семена были с эндоспермом, но при этом редко происходило развитие проростков и редко образовывались у них побеги. В результате скрещивания сорта Пикпуль черный с Ampelopsis acontifolia из 69 частей семян с эндоспермом после 40 дней культивирования на трех вариантах жидких сред выросло только 20 проростков (30%). При этом 17 из них были зелеными, у трех из которых развились побеги (21%). Три проростка были с белыми семядолями и гипокотилями (15% альбиносов от всех развившихся проростков). Из 72 частей семян с эндоспермом скрещивания Пикпуль черный с Parthenocissus inserta развилось меньше как проростков (2 зеленых и 1 белый проросток), так и побегов из них (1 побег).
У всех семян с развившимся эндоспермом, полученных в результате межродовых скрещиваний, в большей или меньшей степени был выражен некроз оболочек семян и эндосперма, что указывает на физиологическую несовместимость родов на биохимическом уровне. Следует заключить, что в дальнейшем необходимо собирать и высаживать семена на более ранних этапах их развития (10, 15 и 25 дней после гибридизации) [1].
Как на жидких средах, так и на твердой среде, на которую были пересажены проростки, наблюдается дальнейшее их аномальное развитие и рост побегов у некоторых из них.
С целью ускорения получения межродовых гибридов между генотипами - представителями различных родов, соцветия которых предварительно за 1725 дней перед цветением, были обработаны колхицином в трех концентрациях - 0,5; 1 и 2%, была проведена межродовая гибридизация. Из обработанных предварительно колхицином соцветий была взята пыльца у видов Ampelopsis acontifolia (0,5; 1 и 2%- ный колхицин), Ampelopsis cordata (0,5 и 1%-ный колхицин) и Parthenocissus inserta (0,5% колхицин) и опылены соцветия у миксоплоидных сортов V. vinifera, соцветия которых также предварительно были обработаны 0,5%-ным раствором колхицина: Харти про Ливье (11 прокастрированных соцветий) и Яхеи (женский тип цветка, 11 соцветий). В качестве контроля 4 соцветия, обработанные 0,5%-ным колхицином, у сорта Яхеи опылены пыльцой Parthenocissus inserta, которая не была обработана колхицином. Из зеленых ягод (25 дней после опыления) выделены незрелые семена, простерелизованы 5%-ным «Доместосом» (10 мин.) промыты стерильной дистиллированной водой и части семян с зародышами (носики семян - после разрезки семян поперек) высажены в жидкую среду Nitsch [14] с добавкой 0,2 мг/л БАП и 0,2 гибберелловой кислоты (ГА3). Всего высажено 857 незрелых семян, полученных в результате указанных выше межродовых скрещиваний. Развившиеся проростки высажены на среду для развития растений in vitro и затем пересажены в теплицу, в условия in vivo.
С целью дальнейшего изучения и выделения истинных межродовых гибридов весной 2009 высажены сеянцы, полученные в культуре незрелых зародышей in vitro. На адаптацию в условиях гидропонной культуры в теплице высажены сеянцы (семена завязались в ягодах, 40 дней после опыления, соцветия развились на побегах, после обработки почек колхицином) скрещиваний Харти про Ливье х Ampelopsis cordata - 2 сеянца в пяти повторностях и Пикпуль черный х Ampelopsis acontifolia - 11 сеянцев в трех-пяти повторностях каждый. Осенью 2009 г. из этих сеянцев получены стандартные саженцы в двух-пяти повторностях. У двух из этих сеянцев сильно рассечены листья (рис.1, 2) (признак рода Ampelopsis), и у одного лоза главного побега окрашена в красный цвет, что свидетельствует о их гибридном межродовом происхождении.

Рис.1. Листья исходных родительских форм Харти про Ливье и Ampelopsis cordata и сеянца, полученного в результате их гибридизации.
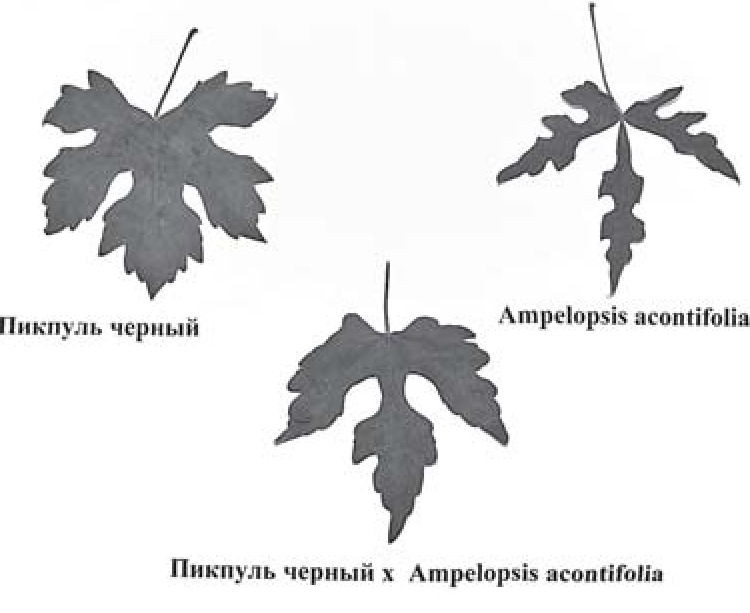
Рис. 2. Листья исходных родительских форм Пикпуль черный и Ampelopsis acontifolia и сеянца, полученного в результате их гибридизации.
С целью получения полиплоидных форм (обработка колхицином распускающихся почек весной 2008 г., контроль - без обработки) для межродовой гибридизации на тетраплоидном уровне в марте 2009 г. в теплицу с гидропонной культурой высеяны семена скрещиваний 2008 г., рассортированные по размерам, следующих генотипов (инцухт, контроль - свободное опыление): Ampelopsis acontifolia (1846 шт.), Ampelopsis cordata (2245 шт.), Ampelopsis serjaniefolia (1723 шт.), Parthenocissus inserta (1162 шт.) и Parthenocissus quinquefolia (701 шт.); у тетраплоида Шасла Гро Куляр белая (313 шт.), и у миксоплоидных сортов рода Vitis вид V. vinifera: Харти про Ливье (1061 шт.), Пикпуль черный (1276 шт.), Шабаш крупноягодный (432 шт.), Шабаш (1244 шт.), Рислинг рейнский (123 шт.) и Мускат александрийский (70 шт.), а также семена, полученные в результате скрещивания Яхеи (женский тип цветка, миксоплоид) х Ampelopsis acontifolia (колхицин) - 34 больших, 26 средних и 55 мелких семян. В конце вегетации, осенью 2009 г., по анатомо-морфологическим признакам полиплоидов [10] и межродовых гибридов отобрано 80 растений для высадки на постоянное место в поле для дальнейшего изучения.
Высажены весной 2009 г. на постоянное место в поле отобранные по морфологическим признакам мощные сеянцы (обработка колхицином распускающихся почек весной 2007 г., выращивание в гидропонной культуре в 2008 г., количество высаженных сеянцев, штук): Parthenocissus inserta - 13, Parthenocissus quinquefolia - 19, Ampelopsis acontifolia - 1, Шасла Гро Куляр белая (инцухт) - 4, Пикпуль черный - 5, Харти про Ливье - 5, Янтарный Магара- ча - 7, Яхеи - 5, Баян ширей (инцухт) - 3, Шабаш (инцухт) - 3, Шабаш крупноягодный - 1, Рислинг рейнский - 10. Также высажены на постоянное место в поле 6 сеянцев (обработка колхицином распускающихся почек весной 2007 г., выращивание зародышей в культуре in vivo и адаптация в гидропонной культуре в 2008 г.) из семян межродового скрещивания Харти про Ливье х Ampelopsis cordata. У одного из этих сеянцев сильно рассечены листья, как и у вида Ampelopsis cordata.
Выводы.
В теплице в гидропонной культуре выращены сеянцы для высадки на постоянное место в поле, полученные в результате межродовой гибридизации (начавшие распускаться почки были обработаны 0,5%-ным колхицином; растения в культуре in vitro были выращены из зародышей выделенных из зеленых ягод - 40 дней после опыления) следующих скрещиваний: Пикпуль черный х Ampelopsis acontifolia (11 сеянцев в трех-пяти повторностях) и Харти про Ливье х Ampelopsis cordata (2 сеянца в четырех повторностях).
Проведено описание по ампелографическим признакам листа исходных форм, относящихся к различным родам семейства Vitaceae, и полученных гибридов, от их скрещивания после полиплоидизации методом колхицинирования.
Поскольку в качестве материнских форм были включены в гибридизацию сорта V. vinifera L. с нерассеченным листом и обладающие функционально женским типом цветка, то можно утверждать, что форму и рассеченность листа гибриды унаследовали от отцовской родительской формы. Следовательно, можно с большой степенью достоверности говорить о получении впервые в мире межродовых гибридов винограда.
Значимость полученных гибридов винограда межродового происхождения заключается не только в том, что это удалось искусственным путем впервые в мире, но что подтверждены биологически обоснованные возможности возникновения таких форм от отдаленных скрещиваний в природе в процессе естественной эволюции.