Во время вегетации параллельно с ростом листьев в них происходят значительные изменения веса сухого вещества и содержания воды, причем процесс направлен к непрерывному увеличению первого и уменьшению второго. Эти изменения графически представляют как бы зеркальное отражение одно другого (Poenaru, Corbeanu, Lazarescu, 1959). В начале вегетации количество воды составляет около 80—82%, в конце вегетации оно уменьшается до 70—75% (в старых листьях падает ниже 70%). В соответствии с этим меняется и содержание сухого вещества в листьях.
Изменяется также общее содержание золы (между 4, 5 и 7,0% с тенденцией к повышению) и минеральных веществ.
По данным Р. Г. Саакян и С. П. Петросяна (1964), количество золы в листьях сорта Гаран дмак колеблется между 6,5—12,3%. Сравнительно более четкое различие в содержании золы установлено по ярусам — в нижнем ярусе золы больше, чем в среднем и верхнем. По мере старения нижних листьев накопление золы увеличивается.
Значительным изменениям подвержено также содержание органических кислот в листьях винограда. По сведениям 3. В. Васильевой (1956), титруемая кислота в листьях разных сортов в течение вегетации непрерывно уменьшается, достигая минимума в период листопада. К. Д. Стоев и И. Димитров (1957) установили, что в период начала созревания ягод в листьях наступает резкое уменьшение общего содержания органических кислот, затем оно снова повышается.
Аналогичная закономерность была выявлена раньше Н. М. Сисакяном, И. А. Егоровым и Б. Л. Африкяном (1948). По их мнению, резкое снижение кислотности в листьях винограда наступает в связи с переходом растения к сахаронакоплению.
Количество хлорофилла и других пигментов также претерпевает значительные изменения: как правило, оно уменьшается со старением листьев.
Изменения в содержании хлорофилла отмечаются также по листьям разных ярусов — больше его в листьях средних зон побега.
На количество пигментов в листьях влияет освещенность и некоторые агротехнические приемы. Наблюдения П. Г. Тавадзе (1957) показали, что по мере ослабления освещенности (до 10—20% от полной) содержание пигментов, особенно хлорофилла, в листьях винограда, как правило, повышается. Увеличивается оно и при повышении влажности почвы до 40—60% от полной влагоемкости. Под влиянием почвенного плодородия, густоты посадки и типа формирования куста содержание пигментов также меняется (Тавадзе, 1952, 1953). Однако после повышения почвенного плодородия путем внесения удобрения и после орошения разница в содержании пигментов под влиянием густоты посадки нивелируется.
Содержание углеводов. В листьях винограда имеется ряд углеводов, которые претерпевают значительные изменения во время вегетации. Кроме крахмала обнаружены сахароза, глюкоза и фруктоза (Стоев, Мамаров, Бенчев, 1960).
Н. М. Сисакян, И. А. Егоров и Б. Л. Африкян (1948) установили, что наибольшее количество крахмала и растворимых сахаров обнаруживается в листьях в фазе начала цветения. В течение вегетации содержание обоих компонентов углеводного комплекса уменьшается и достигает минимума в фазе начала созревания ягод. В фазах полной физиологической зрелости ягод и листопада количество углеводов несколько повышается.
3. В. Васильева (1956), К. Д. Стоев и И. Димитров (1957) показали, что содержание моносахаридов, сахарозы и крахмала увеличивается к концу вегетации. По данным 3. В. Васильевой (1956), максимума количество их достигает к концу первой декады сентября, а по данным К. Д. Стоева и И. Димитрова (1957),—- к периоду листопада (табл. 11).
Таблица 11
Содержание углеводов в листьях винограда (в мг на 1 г абсолютно сухого вещества, в пересчете на глюкозу)
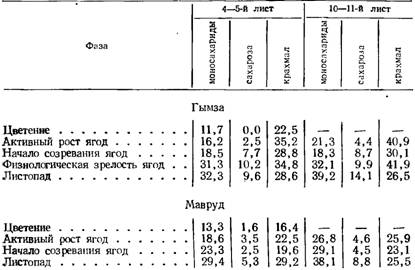
Листья, расположенные в средних ярусах, отличаются более высоким содержанием сахаров, чем листья в зоне грозди. Из растворимых углеводов редуцирующие сахара значительно превалируют над сахарозой (Стоев, Занков, Панайо-тов, Найденова, 1952). По содержанию углеводов листья пасынков не уступают листьями основного побега (табл. 12).
Максимум в содержании углеводов достигается к 10 ч утра, после чего на протяжении дня в количестве их не обнаруживается существенных изменений (Стоев, Занков, Панайо-тов, Найденова, 1952).
Таблица 12
Содержание углеводов в листьях винограда (в % абсолютно сухого вещества)

С. А. Марутян (1957) установила неодинаковое содержание сахаров в листьях плодоносящих и неплодоносящих кустов, что объясняется более интенсивным оттоком пластических веществ у первых. Обнаружены также некоторые особенности углеводного обмена в листьях ранних и поздних сортов. В листьях плодоносящих лоз раннеспелых сортов в большинстве случаев более сильный синтез и гидролиз сахарозы по сравнению с позднеспелыми.
Азотный обмен в листьях винограда. С. А. Марутян (1957) установила, что в листьях позднеспелых сортов больше общего азота и белков, чем в листьях раннеспелых. Это дает основание считать, что между содержанием азотистых веществ в листьях и побегах раннеспелых и позднеспелых сортов обратная связь.
По данным Р. Г. Саакян и С. П. Петросяна (1964), содержание РНК в листьях винограда, составляющее около 4— 5%, к концу вегетации снижается до 1—3%. Не отмечено существенной разницы в содержании РНК в листьях нижнего и среднего яруса. Количество общего и белкового азота в течение вегетации постепенно уменьшается. Распределение азотистых веществ в листьях нижнего, среднего и верхнего ярусов акропетально. Это свидетельствует о том, что синтез белковых веществ интенсивнее протекает в молодых листьях, по мере старения их содержание белкового и общего азота снижается (табл. 13).
Исследования Н. М. Сисакяна, И. А. Егорова и Б. Л. Африкяна (1948) показали, что в ходе вегетации общий и белковый азот в ассимилирующих тканях постепенно уменьшается. При сопоставлении отношения общего и белкового азота выявляется, что перелом в передвижении азотистых веществ в листьях первой формации наступает с фазы начала созревания ягод. В листьях последующей формации сдвиг отношения общего азота к белковому наступает лишь в фазе листопада. Это дает основание считать, что участие ассимилирующих органов различных формаций в создании азотистого баланса неодинаково.
Таблица 13
Изменение содержания азота в листьях винограда (в м г на 1 г веса сухого ацетонового препарата)
| ярус (Л—верхний, Б—средний, В—нижний) | Общий | Белковый | Небелковый |
| До цветения (20/V) |
| |
А | 57,25 | 55,29 | 1,96 |
Б | 50,10 | 48,12 | 1,98 |
В | 48,58 | 46,46 | 2,12 |
| Конец цветения (5/VI) |
| |
А | 49,87 | 48,05 | 1,82 |
Б | 41,63 | 39 98 | 1,65 |
В | 40,15 | 38,65 | 1,50 |
| Формирование и | рост ягод (24/VI) |
|
А | 44,72 | 43,37 | 1,35 |
Б | 37,15 | 35,69 | 1,46 |
В | 31,03 | 29,82 | 1,21 |
| Начало созревания ягод (4/VIII) |
| |
А | 42,40 | 39,76 | 2,74 |
Б | 36,59 | 33,87 | 2,72 |
В | 35,35 | 32,61 | 2,74 |
| Физиологическая | зрелость ягод (5/IX) | |
А | 41,59 | 38,92 | 2,67 |
Б | 34,13 | 32,00 | 2,13 |
В | 35,80 | 33,23 | 2,57 |
К. Д. Стоев, П. Г. Мамаров и И. Б. Бенчев (1960) установили в листьях винограда следующие свободные аминокислоты: аспарагиновую кислоту, глютаминовую кислоту, гистидин, аланин, норвалин, изолейцин, треонин, пролин, тирозин, у-аминомасляную кислоту, валин.
Содержание дубильных веществ и антоцианов. Дубильные вещества принимают важное участие в обмене веществ винограда. Намечается определенная связь между содержанием дубильных веществ в различных органах растения и типом вина.
Н. М. Сисакян, И. А. Егоров и Б. Л. Африкян (1947) изучали изменения дубильных веществ в листьях четырех сортов. Они установили, что по мере старения листьев содержание дубильных веществ претерпевает существенные изменения, причем они имеют некоторую специфичность. У сортов, из которых получают хересные вина (Воскеат и Чилар), обнаруживается два максимума дубильных веществ в листьях: первый в период цветения, а второй в период полной физиологической зрелости ягод. У листьев сортов, которые не хересуются или хересуются плохо, дубильные вещества достигают максимума в период от конца цветения до начала созревания ягод.
К. Д. Стоев и И. Димитров (1957) установили, что в листьях сорта Гымза более нижнего яруса (4—5-й лист) имеются два максимума в содержании дубильных веществ: фаза цветения и фаза физиологической зрелости ягод. У сорта Мавруд второй максимум обнаружен несколько раньше — в начале созревания ягод. В листьях средних ярусов (10—12-й лист) обнаружен лишь один максимум — период с начала созревания до полной физиологической зрелости ягод (табл. 14).
Таблица 14
Динамика дубильных веществ в листьях винограда (в мг на 10 г сухого вещества)

Е. X. Зайцева (1959) обнаружила три максимума в содержании водорастворимой фракции дубильных веществ («собственно танин») в листьях винограда: перед цветением, перед началом созревания ягод и в начале листопада. Полифенол-катехиновая фракция имеет четыре максимума: те же периоды, что и у водорастворимой, и во время технической зрелости ягод; щелочерастворимая фракция — три: после цветения, в период листопада и перед технической зрелостью ягод.
По данным Н. М. Сисакяна, И. А. Егорова и Б. Л. Африкяна (1948), в содержании отдельных фракций дубильных веществ имеются существенные колебания, причем нарастание общего количества танина по мере старения листьев происходит преимущественно за счет полифеиольной фракции.
Количество полифенолов в листьях сортов Воскеат и Мсхали от фазы цветения и до фазы полной физиологической зрелости ягод нарастает, затем до листопада понижается. Содержание флороглюцина не подвергается столь существенным колебаниям в ходе вегетации, как других полифенолов.
С. В. Дурмишидзе (1950) выделил и идентифицировал из дубильного комплекса винограда d-катехин, который находится во всех органах белых и красных сортов. В листьях сорта Саперави в препарате дубильных веществ 15% d-катехина; в растительном материале 0,3%.
В листьях и семенах сорта Саперави С. В. Дуршимидзе (1951) идентифицировал Г-галлокатехин:

Он считает, что дубильные вещества винограда содержат от 45 до 54% галлокатехина.
В других исследованиях С. В. Дурмишидзе и Н. Н. Нуцубидзе (1954) удалось идентифицировать в период полной зрелости ягод в листьях винограда и другие компоненты дубильного комплекса; dl-галлокатехин, dl-катехин и d-эпикатехин-галлат. Таким образом в листьях винограда обнаружено пять различных дубильных веществ: d-катехин; 1-галлокатехин; dl-галлокатехин; dl-катехин; d-эпикатехингаллат.
Дубильные вещества листьев винограда в процессе вегетации претерпевают значительные изменения. В фазу цветения и формирования ягод в листьях имеется лишь dl-галлокатехин; в начале созревания ягод появляется dl-катехин; в период полной зрелости обнаруживаются все пять компонентов дубильных веществ.
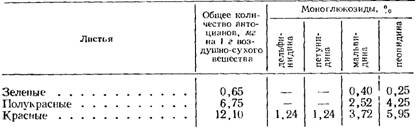
В листьях винограда имеется также ряд антоцианов. Н. Н. Нуцубидзе и Д. И. Гулбани (1959) исследовали листья 50 сортов, выращиваемых в различных почвенно-климатических условиях Грузинской ССР. Они установили, что сорта различаются между собой и по составу антоцианового комплекса и по содержанию отдельных антоцианов (табл. 15).
В листьях одних сортов содержится два, других — три или четыре антоциана моноглюкозидной природы. Сумма глюкозидов мальвидина и пеонидина составляет 70—80% общего количества красящих веществ. Антоциан диглюкозидной природы в красящих веществах листьев винограда встречается как исключение.
При сопоставлении антоцианового состава осенних листьев и кожицы ягод Н. Н. Нуцубидзе и Д. И. Гул бани установили, что красящее вещество листьев, как правило, по составу проще. Состав антоциановых пигментов в кожице ягод и листьях совпадает только у некоторых сортов — Каберне, Саперави, Амурензис и др.
Таблица 15
Состав антоцианов в осенних листьях разных сортов винограда

Антоциановый состав листьев в разные годы часто не совпадает. В течение года содержание антоцианов также значительно изменяется. Неокрашенные и полуокрашеяные листья в основном содержат глюкозиды мальвидина и пеонидина. В период полной окраски листьев обнаруживаются глюкозиды дельфинидина и петунпдина (табл. 16).
Ферментативная активность листьев. Обмен веществ в листьях винограда совершается при участии ряда ферментов. Объектом исследования являлись главным образом ферменты из групп карбогидраз и окислительно-восстановительные.
Активность амилазы и инвертазы в листьях изучали Н. М. Сисакян, И. А. Егоров, Б. Л. Африкян, П. Г. Тавадзе,
Таблица 16
Накопление антоцианов в листьях сорта Саперави
В. П. Соловьева, 3. В. Васильева, К. Д. Стоев, И. Димитров и др.
Н. М. Сисакян, II. А. Егоров и Б. Л. Африкян (1948) установили, что ферментативное образование и распад сахарозы в листовых пластинках в ходе вегетации растения подвергается существенному колебанию. Наибольшая активность ферментативного синтеза сахарозы обнаруживается до начала цветения, после чего наступает ее падение. В последующие фазы развития снова проявляется тенденция к нарастанию синтеза сахарозы. Однако, как отмечают авторы, ферментативный распад сахарозы остается на значительном уровне, что по-видимому, связано с возникновением в растениях оттока веществ из ассимилирующих органов.
П. Г. Тавадзе и В. П. Соловьева (1951) показали, что по фазам вегетации растений активность амилазы в листьях меняется. Во второй половине вегетации они наблюдали усиление гидролитического действия этого фермента, что связывают с оттоком веществ из листьев и накоплением сахаров в ягодах. Гидролитическая активность амилазы сохраняется на высоком уровне и после сбора урожая, во время пожелтения листьев и в период листопада.
Активность амилазы изменяется также в течение суток. Наиболее высока гидролитическая активность в утренние и поздние полуденные, а синтетическая — в полуденные часы.
3. В. Васильева (1956) установила, что активность инвертазы максимальная во время цветения, затем она снижается и достигает минимума в период созревания ягод. В листьях разных сортов она неодинаковая — наиболее высокая у позднеспелых, а наиболее низкая у раннеспелых. На подобную связь между активностью инвертазы и скороспелостью сортов яблони указывают Б. А. Рубин и Н. М. Сисакян (1939).
Между активностью амилазы и инвертазы существует определенная корреляция, на что обратили внимание А. С. Мержаниан и К. Д. Стоев (1948).
К. Д. Стоев и И. Димитров (1957) указали, что активность инвертазы и амилазы неодинаковая в листьях по зонам побега и в различные периоды вегетации: з листьях 4—5-го узла она наиболее высока в мае и июне, причем проявляется при температуре 19—25° С. После начала созревания ягод (вторая половина июля — август) наибольшую активность проявляют инвертаза и амилаза листьев средних ярусов (10—12-й узел), причем при более высокой температуре — 35—42° С. В сентябре активность ферментов выше при относительно низких температурах (25° С). Таким образом, ферментативная деятельность в листьях винограда зависит от внешних условий и в общем следует за изменениями температуры окружающей среды.
По данным 3. В. Васильевой (1956), активность каталазы сравнительно мало изменяется по фазам вегетации. Автор отмечает положительную корреляцию между активностью каталазы и длительностью вегетационного периода: у раннеспелых сортов активность фермента наиболее высокая (сорт Альфа), у позднеспелых (Амурский) — наиболее низкая.
К аналогичному выводу пришли П. Я. Голодрига и Хе Пучао (1963). Они также показали, что разница в активности каталазы у сортов с различной длиной вегетационного периода в наибольшей степени выявляется на ранних фазах развития растений.
По сообщению С. А. Марутян (1957), наибольшая активность каталазы у поздних сортов в начале созревания ягод, а у раннеспелых — в период физиологической зрелости ягод.
Н. В. Новопавловская (1960) установила, что у всех изучаемых ею сортов активность каталазы снижается к концу вегетационного периода.
Значительные различия в активности каталазы обнаружены в листьях по длине побега. Как показали исследования 3. В. Васильевой (1956), Е. X. Зайцевой (1959), Л. В. Колесника (1952), П. Я. Голодриги и Хе Пучао (1963) и др., активность каталазы в нижних листьях значительно ниже, чем в средних и верхних. Пероксидаза, наоборот, активнее в нижних и более старых листьях.
По данным П. Я. Голодриги и Хе Пучао (1963), в поздние фазы развития растений активность каталазы выше у женских сортов. Поэтому они полагают, что можно определить пол растений на ранних этапах селекционного процесса. Пероксидаза активнее у ранних сортов, что ярче проявляется в более ранние фазы вегетации.
С. А. Марутян (1961) считает, что активность пероксидазы в листьях винограда в начале созревания ягод падает, в период физиологической зрелости возрастает. Разницы по активности пероксидазы между ранними и поздними сортами винограда в одни и те же фазы вегетации ей, как и Н. М. Сисакяном, И. А. Егоровым и Б. Л. Африкяном (1948), не установлено.
Активность окислительно-восстановительных ферментов неодинакова и у сортов, различных по устойчивости к морозам. Так, Р. Г. Саакян (1959) установила, что у морозостойких сортов активность каталазы по сравнению с неморозостойкими значительно выше как в начале, так и в конце роста ягод. Она обнаружила также значительно более высокую активность пероксидазы у морозостойких сортов. Активность пероксидазы повышается в течение вегетации, особенно в октябре и с продвижением винограда в горные районы с суровым климатом.
Такую же закономерность наблюдали С. Гребинский (1941), А. Л. Курсанов и Н. Н. Крюкова (1941).
Обнаружена также разница в активности пероксидазы у листьев сеянцев винограда различного пола. По данным С. А. Марутян (1954), в период вегетации пероксидазная активность листьев мужских растений выше, чем женских. У обоеполых растений активность пероксидазы наиболее низкая.
Установлена связь между активностью окислительно-восстановительных ферментов листьев винограда и некоторыми агротехническими приемами. Активность каталазы в листьях перегруженных кустов выше (Библина, 1956; Тавадзе, 1954). Прививка европейских сортов на морозостойкие сорта приводит к повышению активности каталазы в привоях (Новопавловская, 1960). Это влияние наблюдалось в течение всего вегетационного периода и особенно осенью.
Содержание витамина С. В листьях винограда обнаружено некоторое количество аскорбиновой кислоты. По данным С. Я. Мининберг и Н. Н. Фадеевой (1952), оно меняется, причем максимума достигает до цветения и созревания винограда. В листьях раннеспелых сортов в первой половине вегетации (май — август) больше витамина С. В течение суток максимальное количество его обнаружено в 16 ч.
К. Ф. Оинякова (1952) установила, что с 16 июля до 21 августа количество витамина С в листьях сорта Мускат белый увеличивается с 107 мг % До 293 мг %, т. е. в 2— 2,5 раза. Наиболее интенсивно накопление витамина С идет после начала созревания ягод.
Неодинаково содержание витамина С в листьях, находящихся в разных зонах по длине побега: меньше всего в двух нижних листьях.
Значительное различие по содержанию витамина С обнаружено в листьях сортов, неодинаковых по морозостойкости. По данным Р. Г. Саакян (1959), листья морозостойких сортов содержат в 1,5 раз больше аскорбиновой кислоты, чем неморозостойких.
Листья сортов с разной длиной вегетационного периода также содержат неодинаковое количество витамина С. Так, С. А. Марутян (1957) показала, что в листьях поздних сортов его больше.
Н. М. Сисакян, И. А. Егоров и Б. Л. Африкян (1948) установили, что в течение вегетации содержание витамина С в листьях испытывает большие колебания. Как правило, его больше в начальные этапы роста листьев, затем постепенно наступает уменьшение. Сортовых различий в содержании витамина С ими не обнаружено.
Динамика витамина РР (никотиновая кислота) аналогична: наибольшее количество его обнаружено до цветения, наименьшее — в фазе начала созревания ягод.