V.6. КОЛИЧЕСТВЕННЫЕ СВЯЗИ МЕЖДУ ПОГЛОЩЕННОЙ ФАР И ПОКАЗАТЕЛЯМИ ФОТОСИНТЕТИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ. КПД ФАР В ПРОДУКЦИОННОМ ПРОЦЕССЕ ВИНОГРАДНИКА
Основными органами растений, поглощающими лучистую энергию, являются листья, и, естественно, от размеров листовой поверхности зависит количество поглощаемой растением солнечной радиации. В то же время, как показывают исследования, конкретное выражение этой зависимости неодинаково для фитоценозов с различной структурой.
Согласно исследованиям Ничипоровича [251], в посевах однолетних культур с увеличением площади листьев до 30— 40 тыс. м2/га поглощение солнечной радиации непрерывно возрастает (до 75—80% поглощенной ФАР). При дальнейшем росте площади листьев увеличение поглощения происходит медленнее, и при площади листьев около 50 тыс. м2/га кривая поглощения достигает уровня насыщения.
По данным Устенко [380], в посевах кукурузы поглощение ФАР растет по мере увеличения площади листьев и достигает 80% падающей радиации при 60 тыс. м/2га.
Согласно расчетам Каллиса [137], коэффициенты поглощения ФАР фитоценозом зависят от характера размещения листьев в пространстве. Для РП с горизонтальными листьями независимо от времени года получена идентичная зависимость Илф от относительной площади листьев:
LAI........... 1 2 3 4 5 6
аАф........... 0,58 0,79 0,87 0,90 0,91 0,91
При хаотичной ориентации листьев величина аАф изменяется по месяцам и в зависимости от LAI. В летние месяцы в интервале от 23,4 до 50° с. ш. аАф достигает 90% при LAI^5.
Как было показано в главе IV (см. рис. 34), по характеру изменения площади листьев виноградника и поступлению радиации на протяжении вегетации выделяются два качественно различных периода: первый—от начала вегетации до окончания роста побегов, когда одновременно увеличиваются площади листьев и приходы радиации, и второй — от момента окончания роста побегов до уборки урожая, когда площадь листьев сохраняется примерно на одном уровне, а приходы радиации уменьшаются. Следовательно, зависимость коэффициентов поглощения от площади листьев надо было искать для двух указанных периодов.
Из рис. 44 видно, что в интервале исследованных площадей листьев для насаждений с веерными формировками между коэффициентами поглощения и площадью листьев виноградника (тыс. м2/га) обнаруживаются линейные связи, которые для периода роста побегов при ориентации рядов С—Ю аппроксимируются уравнениями:
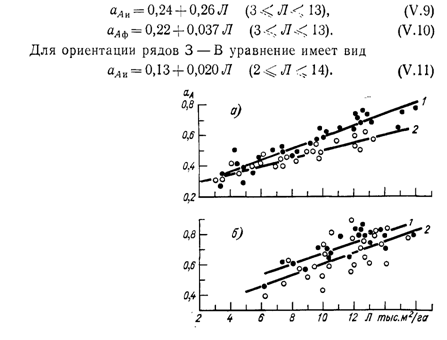
Рис. 44. Зависимость коэффициентов поглощения от площади листьев виноградника при ориентации рядов С—Ю для периода роста побегов (о) и после окончания роста (б) ; — для ФАР, 2 — для ИР.
Для второй половины периода вегетации (рис. 44 б)для насаждений с ориентацией рядов С — Ю аАи/Лаппроксимируется уравнениями:
аАи = 0,26 + 0,034 Л(5 < Л + 16), (V. 12)
аАф = 0.35 + 0,032 Л(5<Л<16). (V.13)
Уравнения связи аАи и аАи позволяют рассчитать поглощение радиации виноградником в период вегетации по данным географических карт распределения солнечной радиации или по данным поступления радиации, полученным метеостанцией. Целесообразно определять поглощение по декадным приходам ФАР. В этом случае по средней величине Лза декаду находят величину аАфи затем по декадному приходу радиации рассчитывают поглощение радиации за декаду. Если кривая роста площади листьев получена за большие, чем декадные, интервалы, то значения средних декадных площадей листьев определяют путем графического дифференцирования. Сумма декадных значений поглощения радиации дает величину поглощения за месяц, за межфазные периоды или за весь период формирования урожая винограда.
Представляло интерес также выявить характер связи между количеством поглощенной ФАР и ЧПФ куста.
Энергетической основой фотосинтеза является энергия ФАР, поглощенная листьями растения. Такая характеристика была получена путем деления суммарного поглощения ФАР кустом (фаза весь период формирования урожая на величину ФП куста — (2дф/ФП. Оказалось, что кусты с однотипными (веерными) формировками в насаждениях с различной густотой посадки характеризуются примерно одинаковыми значениями С>Аф/ФП (табл. 28), за исключением варианта 3 сорта Рислинг, где это соотношение немного меньше. Математическая обработка показала, что в совокупности, представленной вариантами каждого из сортов отдельно, а также в совокупности, представленной вариантами обоих сортов, между величиной <Здф/ФП, с одной стороны, и ЧПФ куста — с другой, коррелятивная связь отсутствует.
Таблица 28
Характеристики поглощенной ФАР единицей площади листьев куста QАф/ФП и средней за вегетацию ЧПФ
Вариант | Сорт Шабаш | Сорт Рислинг | ||||||
QАф/ФП тыс. ккалДм2 день) | ЧПФ | QАф/ФП тыс. ккал/(м2-день) | ЧПФ г/(м2-сутки) | |||||
1970 г. | 1971 г. | 1970 г. | 1971 г. | 1970 г. | 1971 г. | 1970 г. | 1971 г. | |
1 | 1,55 | 1,38 | 3,78 | 4,50 | 2,08 | 1,75 | 5,32 | 6,75 |
2 | 1,70 | 1,35 | 2,74 | 3,87 | 1,80 | 1,64 | 3,48 | 4,18 |
3 | 1,38 | 1,44 | 2,22 | 3,35 | 1,48 | 1,38 | 3,47 | 3,69 |
Примечание. Варианты те же, что в табл. 26.
Сопоставление коэффициентов поглощения с величиной сухой биомассы Мсух в период вегетации показывает, что соотношение между ними неодинаково для виноградников разной густоты посадки: для одной и той же величины биомассы более густые посадки поглощают больше радиации, чем менее густые (рис. 45). Так, например, у сорта Шабаш в 1970 г. в редких посадках на 1 ц сухой биомассы за вегетацию поглощалось 72,6 млн. ккал ИР, а в густых—117 млн. ккал; в 1971 г. соответственно 44 и 68 млн. ккал. Аналогичная закономерность прослеживается и по сорту Рислинг. Следовательно, в насаждениях винограда вес биомассы Убиол не может быть использован для однозначной радиационной характеристики виноградника. Объясняется это тем, что одной и той же величине биомассы может соответствовать разная площадь поглощающей поверхности, которая зависит главным образом от размеров листовой поверхности и ее расположения. В разбираемом случае у обоих сортов единице МСух в густых посадках соответствовала большая площадь листьев (рис. 46).

Рис. 45. Зависимость коэффициентов поглощения ИРаля от величины сухой биомассы МСух в период вегетации при разной густоте посадки.
а — сорт Рислинг, 6 — сорт Шабаш. 1 — 1600, 2 — 6666. 3 — 8888 кустов/га.
Как известно, энергетическая эффективность РП как светопоглощающей системы оценивается по величине КПД поглощенной ФАР (дг)ф). Этот коэффициент представляет собой КПД продукционного процесса и определяется как отношение количества энергии, аккумулированной в биомассе урожая, к количеству поглощенной растениями ФАР за период, в течение которого происходило накопление биомассы [360]:
![]()
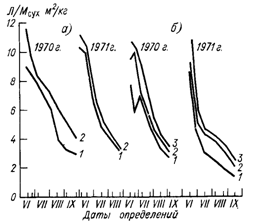
Рис. 46. Изменение соотношения между площадью листьев и сухой биомассой ЛЩсух в период вегетации при разной густоте посадки.
Усл. обозначения см. рис- 45.
![]()
где q — калорийность биомассы (ккал/г); Дт— прирост сухой биомассы РП в интервале времени т; QАф — величина ФАР, поглощенная растительностью за то же время.
Как показано в экспериментальных исследованиях Мурея и Шульгина [226], растения в оптимальных условиях имеют примерно 16-квантовый расход фотосинтеза, при котором КПД ФАР, используемой на истинный фотосинтез, независимо от вида примерно равен 16% у одиночно растущих растений и 19—20% у растений в сомкнутом ценозе. Аналогичные значения, приводимые Тоомингом [360], получены расчетным путем с учетом коэффициента затрат на дыхание. При этом, по [226], КПД ФАР по биомассе для одиночно растущего растения равен 10%, в момент смыкания ценоза —11%, в период максимальных приростов ~12%. Ценоз же в целом имеет КПД по накоплению биомассы ~6% [226].
В естественных, причем оптимальных, условиях КПД, как уже отмечалось, может достигать 4—5% по поглощенной ФАР. В действительности же фактически наблюдаемые коэффициенты значительно ниже и составляют в среднем 0,5—1,5%. В качестве оптимальных принимаются уровни Ау\фв 2—3% [251].
Как показывают исследования, КПД поглощенной ФАР посевов сельскохозяйственных культур изменяется в широких пределах в зависимости от видового состава, структуры ценоза и других факторов. Так, например, в посеве озимой пшеницы величина Ащ изменялась от 1,1 до 6,3% [89]; в посеве хлопчатника она составила 2,2—3,4% [35]; посевы сахарной свеклы при урожае около 1000 ц/га усваивали за сезон 3,4—4,4% поглощенной ФАР, а посевы кукурузы в отдельные периоды — до 6% [97]. Полученные нами данные по поглощенной ФАР и сухой биомассе позволили рассчитать КПД продукционного процесса виноградника в зависимости от его структуры (табл. 29).
Таблица 29
КПД продукционного процесса виноградника дщ (%)
Вариант. Формировка | Сорт Шабаш | Сорт Рислинг | ||||||
Площадь питания куста, м | Аηф по годам | Площадь питания куста, м | Аηф по годам | |||||
1969 | 1970 | 1971 | 1970 | 1971 | 1972 | |||
1. Веерная | 2,5X2,5 | 1,01 | 0,97 | 1,29 | 2,5X2,5 | 1,02 | 1,53 | 1,39 |
2. Веерная | 2,5X1,5 | 0,86 | 0,65 | 1,14 | 1,5X1,0 | 0,76 | 1,04 | 1,16 |
3. Веерная | 1,5X1,0 | — | 0,63 | 0,93 | 1,5X0,75 | 0,93 | 1,07 | 1,33 |
4. Кордонная на Т-образной шпалере | 3,0x1,0 | 1,26 | 0,96 | 1,21 | 3,0x1,0 | 0,89 | 1,71 | 1,33 |
Из табл. 29 видно, что значения Аηф виноградников с различной структурой изменяются в широком интервале (от 0,63 до 1,71%). Сопоставление данных по вариантам с веерной формировкой указывает на тенденцию снижения Аηф по мере увеличения густоты посадки виноградника. Относительно высокими уровнями Аηф в пределах соответствующих лет характеризуются насаждения с кордонными формировками на Т-образной шпалере. У сорта Шабаш значения Ащв этих насаждениях изменяются в интервале от 0,96 до 1,26%; у Рислинга—от 0,89 до 1,71%. По степени использования на фотосинтез поглощенной ФАР насаждения с кордонными формировками на Т-образной шпалере близки к насаждениям с веерной формировкой и редкой посадкой, а в ряде случаев даже выше последних. В целом фактические значения Аηф виноградников (0,86—1,71%) можно характеризовать как средние для агрофитоценоза.
Принимая во внимание приведенные выше данные о соотношении между количеством поглощенной ФАР единицей площади листьев и ЧПФ куста, можно заключить, что снижение Аηф в насаждениях с относительно густой посадкой связано не с обеспеченностью кроны (единицы площади листьев) энергией ФАР, а с эффектом снижения ЧПФ в густых посадках. Иными словами, в густых посадках на единицу листовой (т. е. поглощающей) поверхности производится относительно меньше фотосинтетической продукции, что и ведет к снижению Аηф . В основе же снижения ЧПФ, как мы отмечали, лежат прежде всего не радиационные факторы, а агробиологические, т. е. плодоносность кустов и структура биомассы. Следовательно, повышение показателей Кпд и Кхоз может рассматриваться как один из «внутренних» резервов повышения КПД поглощенной ФАР виноградников, особенно с относительно густой посадкой.