Влияние метеорологических условий на ростовые процессы и накопление биомассы виноградного куста
Биологические аспекты роста зеленых органов растений довольно подробно изучены. Еще в прошлом столетии Сакс установил закон «большого периода роста», согласно которому каждый орган растения или даже какого-либо его участка сначала растет медленно, затем его рост значительно ускоряется, достигает максимума, а затем постепенно замедляется. Объясняется это тем, что на ранних стадиях развития, когда растение еще состоит из одних листьев, все выработанное в процессе фотосинтеза органическое вещество употребляется на создание новых единиц рабочей листовой поверхности, немедленно приступающих к фотосинтезу. В результате общее количество накапливаемого вещества возрастает в геометрической прогрессии. Затем прирост уменьшается и постепенно полностью прекращается, а в конце вегетации нередко происходит даже некоторая потеря сухого веса вследствие того, что листья к тому времени засыхают, процесс фотосинтеза в них прекращается, а трата сухого вещества на дыхание стеблей и плодов еще продолжается.
В настоящее время накоплен определенный опыт по математическому описанию процесса роста и накопления биомассы растений. В. П. Горячкин [16] предложил интегральное уравнение для описания кривой Сакса. А. Демолон [38] считает возможным аппроксимировать данные о росте побегов и листьев винограда по закону сложных процентов, т. е. по уравнению
![]() О) где Р — сухая биомасса в расчетный момент времени, Ро — начальный сухой вес, t —· время, г — мера скорости, е — основание натурального логарифма.
О) где Р — сухая биомасса в расчетный момент времени, Ро — начальный сухой вес, t —· время, г — мера скорости, е — основание натурального логарифма.
Широкое применение в практике лесоводства получило уравнение Е. Бакмана, предложенное им в конце прошлого столетия. Согласно Бакману, характер накопления биомассы лесного фитоценоза можно описать уравнением следующего вида:
![]() (2) где t — возраст, у — прирост в возрасте t, k0, ku k2— постоянные коэффициенты. Это уравнение в литературе получило название функции Бакмана. Ее широко использовал О. С. Вотковский для описания процессов роста леса.
(2) где t — возраст, у — прирост в возрасте t, k0, ku k2— постоянные коэффициенты. Это уравнение в литературе получило название функции Бакмана. Ее широко использовал О. С. Вотковский для описания процессов роста леса.
Особо следует отметить возможность применения функции Бакмана в виноградарстве, поскольку имеются некоторые общие закономерности накопления биомассы у древесных пород и многолетних кустарников или лиан. С. Г. Бондаренко, Т. Я. Кибенко и Н. А. Буякович [16], обработав материал массовых определений прироста биомассы различных сортов винограда, пришли к выводу, что наилучшие результаты расчета накопления биомассы винограда достигаются при приложении для этой цели логистической функции вида
![]() (3) где у — рассчитанное количество биомассы в какой-то период времени, х — время (в днях или декадах), А — максимальное значение биомассы, а и b — коэффициенты, которые находятся методом наименьших квадратов, С — количество биомассы, имеющееся до начала данного периода вегетации.
(3) где у — рассчитанное количество биомассы в какой-то период времени, х — время (в днях или декадах), А — максимальное значение биомассы, а и b — коэффициенты, которые находятся методом наименьших квадратов, С — количество биомассы, имеющееся до начала данного периода вегетации.
Почти такие же результаты авторы получили при аппроксимации экспериментальных данных по параболе и гиперболе второго порядка и биномиальному уравнению, где одна функция степенная, а вторая — экспоненциальная. По нашему мнению, это свидетельствует о том, что процесс роста и накопления биомассы растений можно описать математически различными уравнениями при условии предварительного подбора наиболее удачного вида уравнения для достижения той или иной конкретной задачи.
Характер кривой роста и накопления биомассы растений определяется главным образом характером поступления на зеленую поверхность солнечной энергии. В этом легко убедиться, если построить совмещенные кривые накопления биомассы и сумм эффективных температур или радиационного баланса подстилающей поверхности того места, где изучается накопление биомассы растений (с учетом биологического нуля и длины вегетационного периода данного вида растения) (рис. 2).
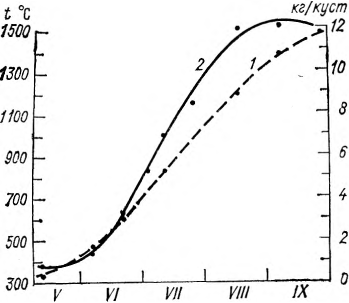
Рис. 2. Кривые нарастания сумм эффективных температур (1) и биомассы винограда Шасла белая (2) (по данным С. Г. Бондаренко). Молдавия, 1973 г.
Наиболее обстоятельные исследования закономерностей накопления биомассы растений винограда проведены С. Г. Бондаренко, Т. Я. Кибенко и Н. А. Буяновичем в Молдавии [14].
Основной вывод, вытекающий из данных этих авторов, убедительно подтверждает S-образный характер роста и накопления биомассы кустов и отдельных органов винограда. При этом максимум накопления биомассы, как и следовало ожидать, зависит от скороспелости сорта (например, у Алиготе на 140-й— 150-й день от распускания почек, а у Саперави— на 160-й—170-й
день). Рост вегетативных органов (побегов, листьев) заканчивается раньше, чем рост гроздей. Максимальный прирост вегетативных органов в зависимости от сорта достигается на 100-й—150-й день вегетации.
Одновременно с сырой биомассой изучалось и накопление сухой массы в органах винограда. Установлено, что вегетативные органы винограда заканчивают накопление сухой массы на 100-й — 120-й день вегетации, а грозди продолжают накапливать сухое вещество и достигают максимума в период полной спелости ягод. Многолетние наблюдения, проводимые нами в Грузии над сортами Ркацители, Цоликаури, Чинури, Цицка и Алиготе, показали, что S-образный рост и накопление биомассы отдельных органов винограда идет по кривой Сакса только в случаях оптимального соотношения тепла и влаги (рис. 3, 4). Нередки случаи, когда условия обеспеченности влагой задерживают рост и в значительной степени искажают характер кривой роста. Поэтому кривые роста и накопления биомассы растений могут быть использованы главным образом при количественном описании общего характера роста. В агроклиматологии они могут применяться при расчетах накопления биомассы растений по многолетним средним данным.
Попытки рассчитать накопление биомассы растений за конкретные годы по этим кривым приводят к существенным погрешностям. Характерным примером может служить рис. 5, на котором изображены кривые накопления биомассы кустов Саперави в Молдавии за 1973, 1974 и 1975 гг.
Согласно рис. 5, кривые в каждом отдельном году сохраняют в какой-то мере S-образный вид, однако количество накопленной биомассы существенно различается. В 1973 г. оно было наибольшим, в 1974 г.— наименьшим, а 1975 г. характеризует почти многолетнее среднее состояние. Таким образом, если вести расчет накопления биомассы по многолетней средней кривой, совпадающей с 1975 г., то в 1973 г. мы получили бы величины, заниженные примерно на 20 %, а в 1974 г., наоборот, завышенные на 30 %.
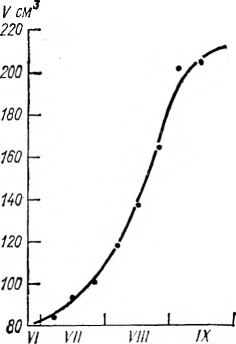
Рис. 3. Динамика роста ягод винограда сорта Ркацители. Дигоми, орошаемые участки, 1969— 1971 гг.
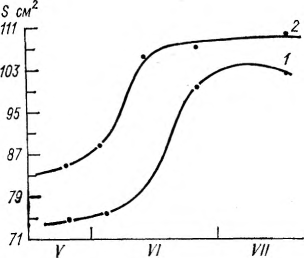
Рис. 4. Динамика роста листьев винограда сорта Ркацители (1) и Чинури (2). Дигоми, орошаемые участки, 1969—1971 гг.
Такие различия в характере кривых роста одних и тех же растений при одинаковой агротехнике обусловлены неодинаково складывающимися агрометеорологическими условиями отдельных лет. Поэтому расчеты нарастания биомассы растений и их отдельных органов необходимо проводить на основе выявления лимитирующих факторов роста и количественной оценки влияния этих факторов на рост.
Воздействие метеорологических факторов на рост растений хорошо изучено. Установлен характер влияния на рост света, температуры, влажности и т. д. В научной литературе по виноградарству встречается много работ, касающихся закономерностей роста побегов винограда [14, 28, 43, 66, 88]. Во всех этих работах подчеркивается, что интенсивность роста побегов винограда в значительной степени зависит от температуры воздуха, а также некоторых других метеорологических факторов.
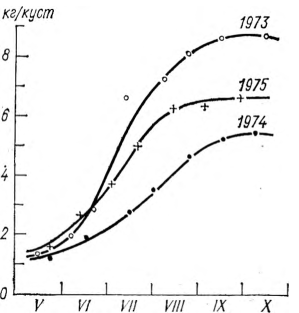
Рис. 5. Динамика накопления биомассы (кг/куст) винограда сорта Саперави. Молдавия, 1973—1975 гг. (по данным С. Г. Бондаренко).
Рост побегов винограда при благоприятных условиях погоды идет очень быстро и может достигать 8—10 см в сутки.
Ученые-виноградари всегда пытались выявить характер влияния факторов внешней среды на интенсивность этих процессов. Французский виноградарь Гильон [28] различает два момента в росте побегов винограда:
- рост верхушечных побегов в точке нарастания;
- рост в длину уже образовавшихся междоузлий.
В первом случае рост идет за счет образования новых клеток образовательных тканей, или прокамбия, во втором случае — за счет удлинения имеющихся клеток древесины и луба. Характер «вытягивания» междоузлий хорошо прослеживается по данным табл. 5, где представлены результаты проведенных Гильоном измерений прироста побегов сорта Фоль-Бланш в Коньяке
Для виноградной лозы, как и для любого другого растения, существуют оптимальные и граничные условия роста, которые определяются биологическими особенностями сортов и состоянием внешней среды. Рост побегов винограда достигает максимума при температурах воздуха 25—35 °С, при понижении температуры до 8°С рост приостанавливается, а при увеличении до 40 °С и выше замечается угнетение роста.
Для изучения зависимости роста побегов винограда от метеорологических условий, выявления лимитирующих рост факторов внешней среды и разработки метода количественной оценки интенсивности роста нами в 1965—1969 гг. были организованы наблюдения в основных районах виноградарства Грузии: в Телави, Болниси и Самгори за сортом Ркацители; в Мухрани — за сортом Чинури; в Сакара и Сочхере — за сортом Цоликаури, а в Ахалцихе — за сортом Алиготе. Прирост побегов измерялся по пятидневкам каждого месяца, начиная со времени распускания почек. Измерения проводились на специально выбранных на каждом участке десяти кустах винограда при общем числе побегов не менее 100. Были использованы также результаты специальных тематических наблюдений за ростом побегов винограда сортов Цоликаури и Цицка, проведенные в 1945—1952 гг. на Сакарской опытной станции виноградарства и виноделия.
Удлинение (мм) виноградных побегов в мае 1903 г. [28]
| Междоузлие | Дни измерений | |||||||
2 | 6 | и | 15 | 18 | 21 | 26 | 30 | |
20-е |
|
|
|
|
|
|
|
|
19 |
|
|
|
|
|
|
|
|
18 |
|
|
|
|
|
|
|
|
17 |
|
|
|
|
|
|
|
|
16 |
|
|
|
|
|
|
| 13 |
15 |
|
|
|
|
|
| 6 | 36 |
14 |
|
|
|
|
|
| 13 | 28 |
13 |
|
|
|
|
|
| 22 | 3 |
12 |
|
|
|
|
|
| 9 | 1 |
11 |
|
|
|
|
| 9 | 2 | 0 |
10 |
|
|
|
| 1 | 9 | 0 | 0 |
9 |
|
| 3 | 2 | 5 | 3 | 0 | 0 |
8 |
| 2 | 2 | 3 | 2 | 6 | 0 | 0 |
7 | 1 | 3 | 2 | 3 | 4 | 7 | 0 | 0 |
6 | 3 | 1 | 3 | 4 | 2 | 2 | 0 | 0 |
5 | 2 | 4 | 6 | 3 | 2 | 0 | 0 | 0 |
4 | 9 | 4 | 1 | 0 | 0 | 0 | 0 | 0 |
3 | 4 | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
1-е | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Показателем интенсивности роста был взят суточный прирост побегов, осредненный по декадам. Оказалось, что побеги винограда наиболее интенсивно растут в период от распускания почек до конца цветения, после чего, несмотря на благоприятные погодные условия, рост ослабевает. Однако на общем фоне этой закономерности в отдельных случаях максимальные значения прироста наблюдаются и после завязывания ягод. Причиной этого, как выяснилось, является угнетенный рост побегов в первый период вегетации, вызванный неблагоприятными погодными условиями.
В табл. 6 и 7 представлены данные об интенсивности роста побегов сортов Ркацители в Телави и Цоликаури в Сачхере в зависимости от погодных условий. Данные эти свидетельствуют о том, что неодинаковая по годам интенсивность роста была вызвана в основном разными условиями увлажнения. Интенсивность роста в 1965—1969 гг. была выше, так как погодные условия первого периода вегетации в эти годы были сравнительно благоприятными: равномерное нарастание температуры воздуха и устойчивое увлажнение. В 1967 г. как температура, так и влажность колебались по декадам в значительных пределах.
Влияние метеорологических условий (по декадам) на интенсивность роста побегов винограда сорта Цоликаури. Сачхере
| Элемент | 1965 г. | 1966 г. | 1967 г. | |||||||||||
июнь | июль | май | июнь | май | июнь | |||||||||
| I | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 2 | 3 | 1 | 2 | 3 |
Средний суточный прирост побегов за декаду, см | 3,1 | 1,8 | 1,2 | 2,3 | 1,2 | 1,6 | 2,2 | 1,1 | 1,8 | 2,6 | 2,8 | 1,5 | 1,7 | 2,1 |
Средняя суточная температура воздуха, ° С | 22,6 | 20,1 | 18,8 | 22,0 | 22,1 | 17,7 | 21,1 | 15,3 | 19,5 | 16,8 | 17,5 | 14,3 | 19,8 | 19,3 |
Сумма осадков, мм | 7,9 | 36,6 | 83,4 | 34,1 | 6,3 | 82,1 | 15,4 | 61,2 | 19,6 | 44,7 | 11,2 | 20,9 | 16,5 | 20,0 |
Число дней с осадками 1 мм | 1 | 4 | 6 | 3 | 1 | 5 | 4 | 7 | 4 | 4 | 5 | 4 | 2 | 3 |
Относительная влажность воздуха в 13 ч, % | 40 | 58 | 64 | 53 | 56 | 48 | 57 | 56 | 49 | 50 | 56 | 48 | 44 | 51 |
Влияние метеорологических условий (по декадам) на интенсивность роста побегов винограда сорта Ркацители. Телави
Элемент | 1965 г. | 1966 г. | 1967 г. | ||||||||||||
май | июнь | май | июнь | май | июнь | июль | |||||||||
| 2 | 3 | 1 | 2 | 1 | 2 | 3 | 1 | 2 | 2 | 3 |
| 2 | 3 | 1 |
Средний суточный прирост побегов, см | 1,9 | 2,3 | 3,0 | 2,1 | 1,7 | 2,3 | 2,6 | 2,7 | 2,2 | 1,5 | 1,2 | 1,1 | 1,1 | 1,9 | 1,1 |
Средняя температура воздуха, ° С | 17,4 | 18,6 | 19,4 | 19,4 | 13,0 | 14,8 | 18,3 | 24,1 | 17,5 | 15,6 | 18,8 | 15,6 | 21,6 | 19,0 | 22,6 |
Минимальная температура воздуха, °С | 11,6 | 14,0 | 16,2 | 14,8 | 9,0 | 10,3 | 13,3 | 18,5 | 13,0 | 11,6 | 13,2 | 10,6 | 15,2 | 14,6 | 17,9 |
Сумма осадков, мм | 22,7 | 39,9 | 23,1 | 65,9 | 25,1 | 69,9 | 20,5 | 18,4 | 21,3 | 26,2 | 12,9 | 8,9 | 27,4 | 27,3 | 3,0 |
Число дней с осадками 1 мм | 2 | 6 | 3 | 5 | 3 | 3 | 3 | 2 | 4 | 2 | 3 | 2 | 1 | 4 | 1 |
Запасы продуктивной влаги, мм | 113 | 117 | 103 | 95 | 94 | 117 | 106 | 85 | 87 | 92 | 53 | 34 | 34 | 59 | 26 |
Относительная влажность воздуха в 13 ч, % | 45 | 59 | 51 | 59 | 55 | 60 | 52 | 45 | 38 | 54 | 45 | 38 | 37 | 49 | 42 |
В связи с этим и рост был неравномерным: во второй декаде мая суточный прирост составил 1,5 см, после чего упал до 1,1 см, а в третьей декаде июня возрос до 1,9 см.
В отдельные годы при неблагоприятных условиях погоды первого периода вегетации рост побегов винограда затягивается и максимальный прирост наблюдается лишь тогда, когда условия погоды этому способствуют. При благоприятных условиях побеги продолжают расти до тех пор, пока не устанавливается оптимальная листовая поверхность, характерная для данного сорта, а также для нагрузки урожаем и системы формировки кустов [148].
Для того чтобы количественно выразить зависимость интенсивности роста побегов от метеорологических условий и этим полнее выявить комплексный характер такой зависимости, проведен корреляционный анализ результатов исследования. Корреляционная связь устанавливалась между двумя, тремя и четырьмя переменными. При подборе коррелянтов, отображающих погодные условия, руководствовались их физиологической значимостью для процессов роста. Таковыми мы сочли, опираясь на литературные данные, среднюю суточную и минимальную температуру воздуха и продуктивные влагозапасы в корнеобитаемом слое, дающие самое объективное представление о влагообеспеченности растений. Показателем интенсивности роста был взят суточный прирост побегов, осредненный по декадам за период распускание почек—конец цветения.
В табл. 8 представлены статистические характеристики связи между вышеуказанными переменными величинами в трех основных виноградарских районах Грузии. Данные эти свидетельствуют о существовании довольно тесной коррелятивной связи между приростом побегов и погодными условиями (коэффициенты множественной корреляции г = 0,86-;- 0,96). Вместе с тем эта связь свидетельствует о комплексном характере влияния погодных условий на рост: из трех факторов, отображающих погодные условия, наиболее тесно связан с ростом тот, который представлен в минимуме. Так, в Телави интенсивность прироста определяется в основном фактором влаги (г = 0,63), тогда как связь прироста со средней суточной и минимальной температурами воздуха представлена соответственно коэффициентами 0,17 и 0,35. В Мухрани же, где виноградники поливаются, самая тесная связь наблюдается между приростом и средней суточной температурой (г=0,89); с запасами влаги эта связь меньше (г = 0,31).
Таблица 8
Статистические характеристики связи между суточным приростом побегов винограда и, средней суточной х и минимальной у температурами воздуха и запасами продуктивной влаги z для основных виноградарских районов Грузии

Особо следует отметить влияние минимальной температуры воздуха на рост. Коэффициенты корреляции между приростом и минимальной температурой возрастают от Телави (0,35) до Мухрани (0,63), так как континентальность климата увеличивается в том же направлении. Следовательно, фактором, лимитирующим рост побегов винограда в предгорных и горных районах, является минимальная температура воздуха.
На общем фоне комплексного характера влияния метеорологических условий на рост в каждом почвенно-климатическом районе один из трех вышеуказанных факторов является лимитирующим. Для богарных районов таким фактором является влажность; в поливных и обеспеченных осадками богарных районах рост лимитируется средней суточной температурой воздуха, а в предгорных и горных районах, как мы уже сказали, — минимальной температурой.
Уравнения регрессии, приведенные в табл. 8, дают приемлемую точность количественной оценки условий роста побегов винограда (10—15%) и могут быть использованы в оперативном обслуживании сельского хозяйства при ежедекадном (ежепентадном) расчете прироста побегов винограда по данным агрометеорологических наблюдений.