Изучение влияния климатических условий на развитие винограда подтвердило, что показатели средних температур воздуха и их суммы недостаточны для обоснования продолжительности некоторых фаз вегетации винограда и для объяснения величины интенсивности сахаронакопления в ягодах винограда. Во многих работах отечественных и зарубежных исследователей это объясняется тем, что для большинства растений умеренных и высоких широт имеет значение не просто средний уровень температуры воздуха, а продолжительность воздействия определенной температуры и характер ее суточного хода.
Работами Е.С. Кузнецовой (1928) и Е.В.Бессоновой (1937) было установлено влияние суточных амплитуд температуры на рост и развитие растений и показано значительное сокращение вегетационного периода многих культурных растений в направлении с запада на восток (на территории европейской части Союза), аналогичного характера были работы А.А.Малышева (1948), З.А.Мищенко (1962). Влияние суточных колебаний температуры на рост и развитие растений впервые было обнаружено и описано А.А. Авакьяном (1936). Результаты его работ подтвердились исследованиями Я.И. Потапенко и Е.И.Захаровой (1940). Наиболее широкие исследования термопериодизма растений были проведены в Калифорнийском технологическом институте Вентом А .А. (1944 ).
Институт "Магарач" занимался вопросом влияния суточного хода температуры воздуха на созревание ягод винограда — на интенсивность сахаронакопления в соке ягод. Тесной связи между величинами средних суточных температур воздуха и средней интенсивности сахаронакопления найти не удается. Она выражена коэффициентом корреляции +0,43 + 0,22. Коэффициент увеличивается до +0,62 + 0,19, если исключить годы с недостатком влаги во время фазы созревания ягод (табл.). Влияние влажности воздуха, продолжительности солнечного сияния, температуры почвы и других показателей на интенсивность сахаронакопления установить не удалось.
Результаты первой попытки найти связь между средней интенсивностью сахаронакопления и суточными амплитудами температуры представлены на рис. 1. Для верхней группы точек коэффициент корреляции + 0,85 + 0,09. Эта зависимость получена для созревания урожая при нормальных климатических условиях (см. таблицу). При недостатке влаги зависимость четко выражена нижней группой точек. Средние температуры периода до некоторой степени связаны с суточными амплитудами температуры. Эта связь значительно ухудшается с уменьшением продолжительности периода. Для средних декадных значений рассматриваемых величин коэффициент корреляции Для августа равен +0,52 +-0.13, для сентября — +0,19 ±0,17.
С 1960 по 1963 г. динамика интенсивности сахаро-накопления исследовалась на фоне микроклиматических наблюдений. Для характеристики температурного режима на участках использовались амплитуды температуры воздуха в центре междурядья на высоте 50 см (рис.2) Аналогичные данные были получены и для других лет. Коэффициент регрессии интенсивности сахаронакопления, зависящий от амплитуды температуры, не остается постоянным во времени. Каждый год он значительно меняется. Это говорит о том, что интенсивность сахаронакопления зависит не только от условий внешней среды в фазу созревания ягод.
Интенсивность увеличения, концентрации сухого вещества в соке ягод - сахаронакопление (Мускат белый) и метеорологические условия
Годы | Периоды | Прирост сахаристости% | Сре-дняя | Сумма температура воздуха, 10° | Сред-няя из суточ-ных ампли- | Сумма осадков, мм | ||||||
за период | сред - ний за 1 сутки | возду-ха за | за период | увеличение сахаристости на 1% | за месяц до нача-ла пе- риода | за 1-ю | за | |||||
1947 | 25/YIII-20/IX | 6,9 | 0,27 | 20,0 | 321 | 75 | 6,3 | 13,8 | 2,3 | 1,1 | ||
1948 | 20/YIII-30/1Х | 13,1 | 0,32 | 19,9 | 816 | 62 | 6,4 | 13,1 | 32,2 | 1,2 | ||
1949 | 20/YIII-16/1Х | 7,1 | 0,10 | 20,8 | 769 | 108 | 6,6 | 0 | 6,3 | 42,2 | ||
1950 | 15/YIII-20/1Х | 11,3 | 0,31 | 22,6 | 813 | 72 | 6,9 | 17,5 | 3,0 | 4,7 | ||
1951 | 15//Ш-20/1Х | 12,0 | 0,33 | 24,0 | 863 | 72 | 6,6 | 10,5 | 23,9 | 0 | ||
1952 | 15/YIII-20/1Х | 11,1 | 0,31 | 23,8 | 857 | 77 | 6,7 | 20,7 | 3,3 | 6,6 | ||
1953 | 15/YIII-20/1Х | 12,5 | 0,35 | 22,0 | 793 | 63 | 7,2 | 2,4 | 20,1 | 10,3 | ||
Годы | Периоды | Прирост сахаристости % | Средняя температура воздуха за период | Сумма температур воздуха, 10°С | Средняя из су- точный | Сумма осадков, мм | |||||||||||
за период | за 1 сутки | за период | уве | амплитуд температурь воздуха за период | за месяц | за | за | ||||||||||
1954 | 15/YIII-20/1Х | 14,8 | 0,41 | 24,4 | 880 | 59 | 7,7 | 12,8 | 2,0 | 7,2 | |||||||
1955 | 15/YIII-20/1Х | 13,1 | 0,36 | 23,4 | 844 | 34 | 7,0 | 40,1 | 21,7 | 0 | |||||||
1957 | 25/YIII-25/1Х | 6,8 | 0,22 | 22,7 | 705 | 104 | 6,9 | 1,7 | 1,3 | 2,8 | |||||||
1958 | 9/YIII-11/IX | 12,4 | 0,38 | 22,0 | 727 | 59 | 7,2 | 13,7 | 0* | 39,7 | |||||||
1959 | 20/YIII-17/IX | 8,3 | 0,30 | 20,8 | 583 | 70 | 7,0 | 26,2 | 19,7 | 23 | |||||||
1960 | 19/YIII-16/1Х | 8,2 | 0,29 | 20,4 | 572 | 70 | 6,3 | 78,4 | 27,8 | 3,8 | |||||||
1961 | 9/YIII-20/ IX | 11.7 | 0,28 | 22,5 | 954 | 82 | 7,6 | 4,0 | 0,5 | 13,5 | |||||||
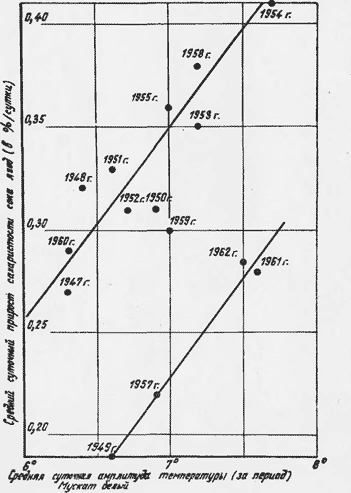
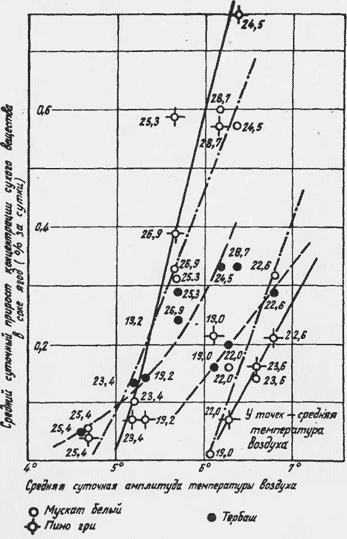
Рис. 2. Средний суточный прирост концентрации сухого вещества в соке ягод и суточные амплитуды температуры воздуха (1962 г.).
Она зависит и от потенциальных возможностей виноградного куста, сложившихся в результате климатических условий в предшествующие фазы вегетации, от условий питания, формирования куста, нагрузки и других факторов. Судя по полученным материалам, сахаронакопление возможно только при амплитудах, превышающих определенное их значение. Этот порог не остается постоянным, он меняется в зависимости от сорта винограда и условий обеспеченности винограда влагой. Во время засушливых периодов он больше. Минимальное значение порога, очевидно, имеет значение 4-5 . Увеличение амплитуд сказывается на интенсивности сахаронакопления только до какого-то их значения, после чего реакция исчезает.
На рис.3 видно увеличение интенсивности сахаронакопления с повышением температуры. С I960 по 1963 г. наибольшая из средних температур семидневного периода (через такие промежутки времени определялась концентрация сухого вещества в соке ягод) достигала 28,7 . До этих температур воздуха депрессия интенсивности сахаронакопления не отмечалась (Мускат белый, Пино-гри, Тербаш).
Показатель суммы активных температур воздуха, рассчитанных на 1% увеличения сахаристости во время различных периодов, очень изменчив (см.табл. ).В те периоды, когда виноград плохо обеспечен влагой (1949, 1957, 1961 гг.), эти суммы особенно велики.
Но и в остальные годы суммы изменчивы (59-77 ). Данные, приведенные в таблице, позволяют сделать вы вод: сумма температур, рассчитанная на 1% увеличения концентрации сухого вещества в соке ягод, имеет тенденцию увеличиваться с уменьшением интенсивности сахаронакопления. Они тем больше, чем больше нужно времени для изменения сахаристости на 1%.
Если интенсивность сахаронакопления имеет прямую корреляцию с суточными амплитудами температур.
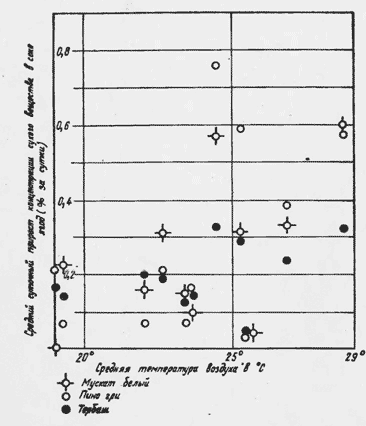
Рис. 3. Средний суточный прирост концентрации сухого вещества в соке ягод и средние температуры воздуха (1962 г.).
Одна из попыток найти зависимость между продолжительностью межфазового периода от начала созревания ягод до их промышленной зрелости для сорта Халили белый представлена на рис. 4.
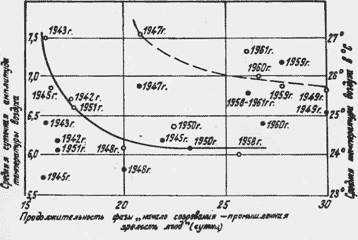
Рис.4. Амплитуда температуры воздуха, средние температуры и продолжительность фазы "начало созревания” — промышленная зрелость ягод у винограда сорта Халили белый.
Расположение точек позволяет выразить зависимость двумя кривыми : для нормальных и для неблагоприятных условий (недостаток влаги или сочетание недостатка влаги с высокими температурами). На этом же рисунке представлена продолжительность периода при различных средних температурах воздуха. В этом случае расположение точек не позволяет установить влияния температуры на продолжительность межфазового периода. Здесь даже намечается тенденция увеличения продолжительности его с повышением температуры для всего диапазона наблюдавшихся температур.
Несомненно, суточные колебания температуры воздуха, обусловленные особенностями рельефа местности
должны учитываться при выборе участков под насаждения ценных сортов винограда. Суточный ход температуры воздуха должен предусматриваться и при сорторайонировании, поскольку сглаженный суточный ход задерживает созревание урожая.
Основным методом, применяемым при изучении количественного содержания углеводов и других соединений в органах виноградного растения, является метод биохимического анализа. Результаты анализа выражаются в процентах к сухому веществу. Однако, с биологической точки зрения, данные обычного биохимического анализа лишь при строго ограниченных условиях могут рассматриваться как достаточно точные и сравнимые, так как они не всегда позволяют выявить действительные изменения количества различных веществ в клетках тканей и органов растения.
Описываемый ниже метод дает возможность установить абсолютное количество тех или иных веществ, приходящихся в среднем на одну клетку изучаемого растительного материала. Сущность метода состоит в обработке данных биохимического анализа с учетом результатов микрометрических измерений, позволяющие определить число клеток в анализируемых образцах.
Пробы растительного материала отбирают в виде сегментов изучаемых органов или же в зависимости от целей исследования частей органов и отдельных тканей. Образны разделяют на две группы, представляющие средние пробы изучаемого материала. Первая группа служит для измерения их размеров (высота, длина, ширина, радиус или диаметр и т.д.), определения среднего веса клетки и для биохимических анализов; вторая — для определения числа клеток в отобранных образцах. Эту группу образцов можно использовать как в свежем виде, так и после консервирования их способами, не вызывающими изменения размеров анатомических элементов (для виноградного растения с этой целью может применяться смесь равных частей этилового спирта, глицерина, воды и 1-процентного формалина). С помощью окуляр-микрометра или иного микрометрического устройства подсчитывают на продольных (радиальных и тангентальных) и поперечных анатомических срезах под микроскопом количество клеток, приходящееся на линейную единицу по различным сечениям образца.
Пользуясь полученными данными, определяют число клеток, входящее в состав образцов первой группы, по формуле:
![]()
где l, В, Н — длина, ширина и высота образца;
м,п,р — среднее число- клеток на линейную единицу по длине, ширине и высоте образца.
Все образцы первой группы или среднюю пробу из них высушивают и определяют средний вес абсолютно сухого ( сырого) -вещества одной клетки по формуле:
![]()
где O - вес образца в абсолютно сухом (сыром ) состоянии ; а - число клеток, составляющих образец.
По аналогичной схеме может быть определен средний вес сырого вещества одной клетки растительных органов и тканей. Для этого образцы первой группы взвешивают не в абсолютно сухом, а в свежем состоянии
Количество отобранных образцов и повторностей измерений должно обеспечивать достаточную статистическую достоверность полученных данных, определяемую на основе вариационного анализа.
Образцы первой группы измельчают и используют для биохимических анализов на искомые соединения. Анализы выполняют по методике, позволяющей получить наиболее точные результаты, выраженные в процентах к весу сухого или сырого вещества навески.
Пользуясь данными биохимического анализа и определения среднего веса клетки, находят абсолютное количество искомого соединения, приходящегося на одну клетку, по формуле:
![]()
где а — средний вес абсолютно сухого (или сырого) вещества одной клетки;
д - содержание искомого соединения в процентах к сухому или сырому веществу.
Приведенные формулы (1,2,3) могут быть изменены в зависимости от изучаемых объектов, задач исследования и способов расчета.
Для анализа междоузлий виноградных побегов можно применить следующую методику.
Тщательно измеряют свежесрезанные сегменты побегов, взятые на определенных междоузлиях А, после чего их высушивают, взвешивают, измельчают и подвергают биохимическому а на лис у. Отрезки тех же междоузлий, примыкающие к торцовым частям взятых сегментов Б (рис.1), консервируют в жидкости, не вызывающей изменения размеров анатомических элементов. Путем микрометрических измерений на полученных тангентальных и радиальных срезах, толщиной 10-15 мм, подсчитывают число клеток, приходящееся на объем отрезка, соответствующий определенной высоте. Дли цилиндрических форм побега число "а" составляющих их клеток может быть вычислено по формуле:
![]()
где в — среднее число клеток по радиусу отрезка;
с - среднее число клеток по его высоте.
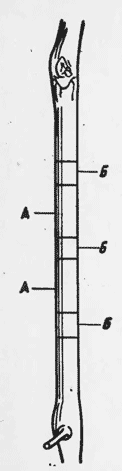
Рис.1. Отбор образцов виноградного побега для биохимических анализов (А) и микрометрических измерений (Б).
(4)
Так как побег винограда имеет дорзовентральное строение и поперечник его по разным направлениям неодинаков, формула (4) может быть использована только при условии приведения частных значений поперечника к среднему диаметру цилиндра. Для этого на поперечных срезах побега через равные угловые промежутки в радиальным направлении измеряют расстояния от центра сердцевины до внешней границы коры, подсчитывают число приходящихся на каждое направление клеток и из полученных данных выводят средние показатели.
Для контроля можно подсчитать с помощью окулярной микрометрической сетки число клеток, приходящееся на единицу площади поперечного сечения, найденную по среднему радиусу среза. В этом случае применима формула:
![]()
где Т - число клеток, приходящееся на плоскость поперечного сечения среза побега.
Подсчет числа клеток, приходящегося на линейную единицу отрезка по его высоте (вдоль оси побега), выполняют на тангентальных срезах, сделанных через равные расстояния, путем последовательного наложения микрометрической линейки (А.А.Яненко-Хмелевский, 1854).
Дальнейшую работу, связанную с определением среднего веса одной клетки в анализируемых образцах, выполняют, используя биохимические анализы и перерасчет аналитических данных на одну клетку (как описано выше).
Различия между показателями процентного содержания соединений и абсолютного количества их в клетках имеют важное значение. На основе тех и других данных могут быть сделаны диаметрально противоположные выводы. Об этом свидетельствует приводимый ниже пример анализа на содержание лигнина в междоузлиях побегов у сорта Сенсо.
Результаты определения среднего веса сухого вещества одной клетки в 8-10-м междоузлиях побегов винограда показывают, что в течение вегетации накопление веществ в тканях побега происходит более интенсивно, чем их трата, и средний вес абсолютно сухого вещества одной клетки все более увеличивается. За период с 15 мая по 10 октября отмечено увеличение сухого вещества одной клетки от 0,0136 до 0,0366 мкг (рис.2).
Определение количества лигнина в тех же объектах проведено по общепринятым методам биохимического анализа и по описанному способу (рис.3). На рисунке масштаб шкал выбран так, чтобы показатели содержания лигнина, полученные разными способами 15 мая (первый срок определений), находились на слизком уровне.
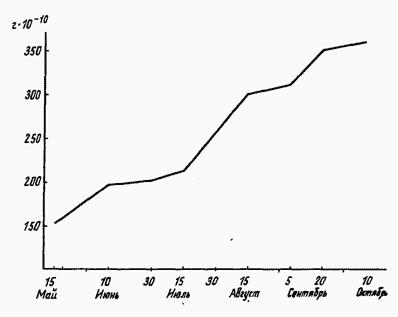
Рис. 2. Изменения среднего веса сухого вещества одной клетки побега на протяжении вегетации. Сорт Сенсо.
С изменением веса клеток изменяется абсолютный вес приходящихся в среднем на одну клетку анализируемых соединений. Кривая показывающая динамику абсолютного количества лигнина, приходящегося на одну клетку, резко отличается от кривой 2, отражающей ход процентного содержания лигнина. Вследствие прогрессирующего возрастания среднего веса сухого вещества .одной клетки побега ход фактического накоплений клетками лигнина все более отклоняется от хода содержания его, выраженного в процентах к сухому веществу, и в конце вегетации разрыв между показателями, полученными с помощью обоих методов, достигает максимальной величины.
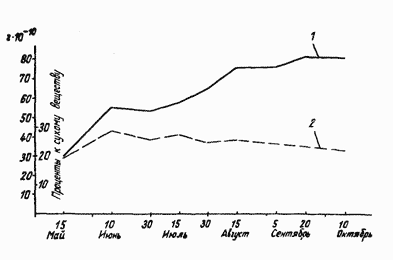
Рис. 3. Динамика содержания лигнина в побегах винограда: 1-вес, приходящийся на одну клетку; 2 - в процентах к сухому веществу.
Полученные данные приводят к выводу, что наблюдаемое во второй половине вегетации систематическое снижение процентного отношения лигнина к сухому весу тканей побега не может рассматриваться как уменьшение его фактического количества, так как абсолютное содержание лигнина в клетках побега продолжает, хотя и слабо, возрастать либо остается примерно на одном уровне. А так как одновременно происходит значительное увеличение сухого веса клеток, преимущественно вследствие накопления пластических соединений, то этот процесс при некритической оценке результатов биохимического анализа может рассматриваться как уменьшение количества лигнина.