В этом разделе описана совокупность превращений сахаров Сб производимых дрожжами, известная под названием спиртового брожения. Однако в строгом смысле слова спиртовое брожение означает исключительно образование спирта из сахаров согласно уравнению Гей-Люссака С6Н1206*2СН3 - СН20Н+-2С02.
Но известно, что во время развития дрожжей, например, в бродильном чане не все молекулы сахара подвергаются такому превращению; некоторое число их разлагается в процессе глицерино-пировиноградного брожения согласно уравнению Нейберга
С6Н1206-<- СН2ОН - СНОН - СН2ОН +
Глицерин
+ СН3-СО — СООН.
Пировиноградная кислота
Наряду с глицерином образуется пировиноградная кислота, в некоторых случаях декарбокоилированная в этаналь, но не восстановленная в спирт; она представляет собой основу для образования разнообразных вторичных продуктов.
Спиртовое брожение
Гликолиз является первым химическим актом спиртового брожения. Образующаяся пировиноградная кислота декарбоксилируется в ацетальдегид и восстанавливается в этиловый спирт. Эта реакция реализуется окисленной формой NAD, которая появляется в процессе окисления глицерин- альдегида-3-фосфата, следовательно, обе соответствующие реакции являются спаренными; они представляют окислительно-восстановительный процесс (рис. 5.12). Необходимость вторичного окисления NADH2 понятна. Если бы его не было, гликолиз прекратился бы с того момента, когда весь NAD, присутствующий в клетке, был бы восстановлен.
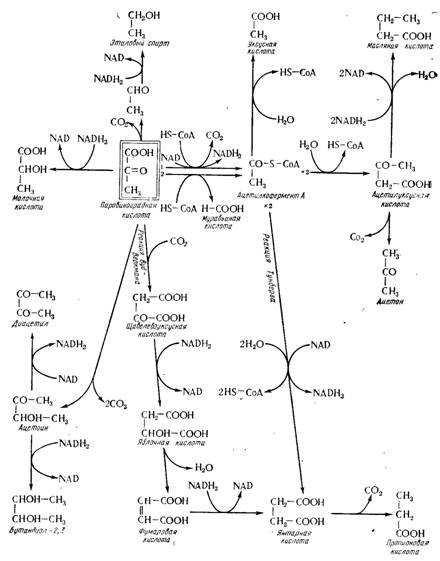
Рис. 15.11. Эволюция пировиноградной кислоты в анаэробиозе.
Энергетический баланс спиртового брожения идентичен балансу гликолиза, т. е. на одну молекулу разложенного сахара образуется 2 молекулы АТФ. Химический баланс брожения, осуществляемого дрожжами, можно записать следующим образом: c6hi206+ 2адр + 2н3ро4 2сн3 - сн2 0н+ + 2с02 + 2атр + 2н20.
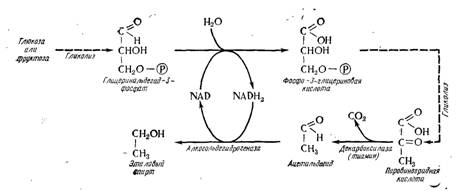
Рис. 15.12. Схема спиртового брожения.
От химического превращения молекулы глюкозы в С02 и этанол выделяется 40 ккал (Конн и Штумпф, 1972); поскольку энергия образования связи АТФ равна 7,3 ккал, из высвобожденных 40 ккал 14,6 используются дрожжевыми клетками для обеспечения их жизненных функций, в частности размножения. Разница, т. е. 25,4 ккал, высвобождается в виде теплоты и вызывает нагревание бродящей среды.
Разложение гексоз в аэробиозе во время дыхания происходит в соответствии с классическим уравнением: С6Н!206 + 602 6С02 + 6Н20.
Вторичное окисление NADH2 за счет кислорода воздуха, с одной стороны, и окисление пировиноградной кислоты при посредстве цикла Кребса, с другой, обеспечивают восстановление значительно большей энергии, соответствующей 38 молекулам АТФ на одну молекулу сахара. Соответствующая энергия выделяется из свободной энергии химического уравнения дыхания, которая составляет 686 ккал на одну молекулу сахара. Отсюда понятно, когда говорят, что «брожение — это плохое использование энергии углеводного питания», это значит, что «небольшое число дрожжей разлагают большое количество сахара».
Но сбраживание сахаров дрожжами всегда ведет к появлению небольшого количества молочной кислоты, порядка 400 мг/л, соответствующего превращению 0,0005% молекул сахара (Пейно и сотрудники, 1967 6).
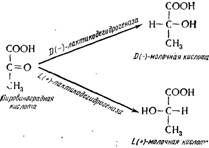
Рис. 15.13. Образование двух изомеров молочной кислоты из пировиноградной кислоты.
Механизм этого образования близок к гомоферментативному брожению (см. рис. 15.25); пировиноградная кислота не подвергается действию двкарбоксилазы, чтобы быть восстановленной непосредственно лактодегидрогеназой в молочную кислоту (см. рис. 15.11). Но существуют два изомера молочной кислоты [молочная кислота D (—) и L (+)], которые образуются из пировиноградной кислоты соответствующими дегидроген азами (рис. 15.13). Дрожжи образуют почти исключительно молочную кислоту D (—); исключением является Saccharomyces veronae, так как они обладают специфическим свойством образовывать только молочную кислоту L (+)> во всех случаях намного больше любого вида (Пейно и сотрудники, 1967 а).
Глицерино-пировиноградное брожение
При описании механизмов спиртового брожения авторы показали необходимость вторичного окисления NAD, восстановленного во время гликолиза, причем это окисление происходит за счет ацетальдегида. Но в начале брожения ацетальдегида в среде нет. Для того чтобы гликолиз протекал нормально, необходимо присутствие ацетальдегида, но он образуется только в результате гликолиза.

Рис. 15.14. Схема глицерино-пировиноградного брежения.
Таким образом объясняется существование своего рода пускового периода, периода индукции, в течение которого повторное окисление NADH2 происходит за счет молекулы дигидроксиацетонфосфата, которая сама образуется при разрыве молекулы сахара в процессе гликолиза (см. рис. 15.9). Это восстановление ведет к образованию глицерина (рис. 15.14). Одновременно фосфо-З-глицериновая кислота, получающаяся от окисления глицер и н а л ь дегида -3 - фосфата, разлагается согласно реакциям гликолиза на пировиноградную кислоту, которая сама в дальнейшем декарбоксилируется в ацетальдегид. Но этот ацетальдегид не может быть восстановлен в спирт потому, что два атома водорода (происходящие из NADH2), которые обеспечивают это восстановление в случае спиртового брожения, уже были использованы для образования глицерина из дигидроксиацетонфосфата. Каждый раз, когда происходит образование одной молекулы глицерина, одна молекула пировиноградной кислоты (или ацетальдегида) накапливается без превращения в молочную кислоту (или в этиловый спирт). Ниже будет показано, что эта пировиноградная кислота, получающаяся в результате глицерино-пировиноградного брожения, составляет основу многих вторичных продуктов.
Сопоставление схем спиртового и глицерино- пировиноградного брожения (см. рис. 15.12 и 15.14) показывает, что существует антагонизм между двумя акцепторами водорода, ацетальдегидом, с одной стороны, и дигидроксиацетонфосфатом, с другой. Поскольку первый легче восстанавливается, именно он и фиксирует два атома водорода, происходящие из NADH2, когда эти два вещества находятся в эквивалентных концентрациях. Но если ацетальдегид отсутствует, а именно такое положение и имеется в начале развития дрожжей в среде, содержащей сахар, дигидроксиацетонфосфат служит акцептором водорода и в то же время обеспечивает накапливание Ацетальдегида, необходимого для возбуждения спиртового брожения в собственном смысле слова.
В целом механизмы спиртового и глицерино-пировиноградного брожения тесно связаны между собой. В начале развития дрожжей преобладает глицерино-пировиноградное брожение и, как следствие, образование глицерина, но даже в период бурного брожения никогда не бывает чисто спиртового брожения.
Учитывая тот факт, что в вине содержится приблизительно 8 г/л глицерина, или 0,087 молекулы, происходящей из 0,087 молекулы сахара на 100 г этанола, или 2,2 молекулы, происходящие из 1,1 молекулы сахара, можно подсчитать, что при брожении вина около 8% молекул сахара идет на глицерино-пировиноградное брожение и 92%—на собственно спиртовое брожение.
Вторичные продукты, образующиеся из пировиноградной кислоты
Как было указано выше, пировиноградная кислота, получающаяся в результате глицерино-пировиноградного брожения, не может быть восстановлена в молочную кислоту или в этаналь после декарбоксилирования, потому что атомы водорода, необходимые для этого восстановления, уже использованы для образования глицерина.
В конце брожения в вине находится лишь небольшая часть пировиноградной кислоты, поскольку вино всегда содержит ее в малых количествах (в среднем 80 мг/л, Блуэн и Пейно, 1963). Но она идет прежде всего на образование различных вторичных продуктов в соответствии с механизмами, общими для многих видов брожения (см. рис. 15Л1).
Сначала переход в ацетилкофермент А обеспечивает образование уксусной кислоты, которая обеспечивает всякое брожение; первый этап этой реакции может протекать на основе двух механизмов, один из которых включает окислительное декарбоксилирование, другой — образование муравьиной кислоты. Первый, конечно, более важен потому, что производство муравьиной кислоты всегда очень незначительно, несколько граммов на 1 л (Гимберто, 1969), частично этерифицированной (Бертран и Риберо-Гайон, 1968). Исходя из роли ацетилко- фермента А, можно также объяснить образование масляной кислоты и ацетоина в вине в виде следов, первая — в виде эфира.
Также на основе пировиноградной кислоты конденсация двух молекул, сопровождаемая декарбоксилированием, ведет к ацетоину или ацетилметилкарбинолу, который может дать сам или бутандиол-2,3 путем восстановления, или диацетил путем окисления; эти ацетоиновые вещества находятся в вине.
Можно также иметь в виду карбоксилирование пировиноградной кислоты (реакция Вуд-Веркмана), ведущее к образованию щавелево-уксусной кислоты. Эта кислота может быть превращена в яблочную кислоту, образование которой показали Карль и сотрудники (1967); но в брожении вина
Это превращение маскируется разложением яблочной кислоты натурального виноградного сока. Наконец, яблочная кислота дает через посредство фумаровой кислоты янтарную кислоту, о которой со времен Пастера известно, что она представляет один из важнейших вторичных продуктов спиртового брожения. Этот же механизм ведет также к пропионовой кислоте, присутствующей в вине в виде следов, в частности в форме сложного эфира (Бертран и Риберо-Гайон, 1968).
Но янтарная кислота может быть также образована конденсацией двух молекул ацегилкофермента А (реакция Тунберга). Трудно определить, который из двух возможных механизмов ответствен за образование этой кислоты, так как имеются аргументы в пользу и того и другого: возможно, что они функционируют в дрожжах оба одновременно.
К продуктам брожения, образующимся из пировиноградной кислоты (см. рис. 15.11), нужно добавить лимонно-яблочную кислоту, а также диметилглицериновую кислоту. Лимонно- яблочную кислоту впервые идентифицировали Карль и сотрудники, ее концентрация в вине находится в пределах от 0 до 300 мг/л (Дюбюс, 1969). Карль и сотрудники предложили гипотезу об образовании лимонной кислоты в процессе бактериального брожения согласно реакции, напоминающей яблочно-молочное брожение (рис. 15.15). Но этот механизм не подтвердил Димотаки-Кураку (1962), который показал, что уменьшение содержания лимонной кислоты во время яблочно-молочного брожения не сопровождается образованием лимонно- яблочной кислоты. Последняя появляется исключительно в процессе спиртового брожения, вызываемого дрожжами. Елшин (1962) показал на высших растениях образование лимонно- яблочной кислоты при конденсации уксусной кислоты в виде ацетилко- фермента А с Пировиноградной кислотой (см. рис. 15.15); можно предположить функционирование такой реакции в дрожжах.
Имеются сообщения об образовании лимонной кислоты в ходе спиртового брожения. В действительности методом количественного химического анализа, использованного этим автором, определяли одновременно и лимонную, и лимонно-яблочную кислоту. Сейчас известно, что образуется только последняя, а первая остается без изменения.
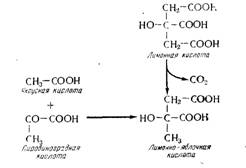
Рис. 15.15. Механизм образования лимонно-яблочной кислоты.

Рис. 15.16. Диметилглицериновая кислота.
Диметилглицериновая кислота (рис. 15.16) была идентифицирована в вине Карлем и сотрудниками (1966); ее концентрация находится в пределах от 0 до 600 мг/л (Дюбюс, 1969). Карль и сотрудники (1967) предполагают, что она также образуется из Пировиноградной и уксусной кислот согласно реакции, родственной той, которая схематически показана на рис. 15.15.