М. К. КАРАЕВ
Дагестанская государственная сельскохозяйственная академия
Продуктивность виноградного растения зависит от работы листового ассимиляционного аппарата, использующего солнечную энергию на создание органической массы растений, в том числе и хозяйственно важную ее часть — урожай винограда. В связи с этим для получения высоких качественных урожаев необходимо, прежде всего, обеспечить непрерывное с начала вегетации максимально возможное развитие активной в фотосинтезе ассимиляционной поверхности растения. Очень большое значение при этом имеет характер расположения листового аппарата. Повышение урожая и его качества обусловлено способами ведения и формирования кустов, позволяющими улучшать освещенность листового аппарата на фоне высокой агротехники и повышать использование солнечной энергии. С этой точки зрения несомненный интерес представляют насаждения с длиннорукавными формами кустов на одно- и двухпроволочной шпалере при свободном развитии побегов, более рационально использующими воздушно-световое пространство и отличающимися повышенной продуктивностью, для практики. По нашим данным, при свободном ведении прироста ослаблялось проявление полярности, а это приводило к подавлению ростовых процессов и уменьшению облиственности побегов.
Каждому растению на шпалере предоставляется определенное пространство, в котором ассимиляционный аппарат должен располагаться наиболее рационально без загущения и максимально использовать окружающую среду для фотосинтеза. Достигается это прежде всего формой куста, характеризующейся наличием определенных структурных элементов и их размещением в пространстве. Это, в свою очередь, создает определенный режим освещенности и фитоклимата. В связи с этим выяснение вопроса продуктивности фотосинтеза листьев исследуемых форм куста представляет определенный интерес.
Для количественной оценки пространственного распределения элементов фитомассы используют различные фотометрические характеристики, к наиболее простым из которых относятся параметры кроны (ширина, длина, высота), число побегов на кусте и на одном погонном метре ряда, длина одного побега и его облиственность, характер распределения площади листовой поверхности относительно горизонтальной (JLAJг) и вертикальной (JLAJв) поверхности проекции кроны, плотность ее в кроне куста (А. Г. Амирджанов, 1980).
Согласно нашим исследованиям крона кустов в зависимости от формы имеет разную архитектуру. При одинаковой длине кроны в зависимости от способа ведения прироста и формирования куста меняются его ширина и высота.
Так как схема посадки в наших исследованиях была одинаковой для всех форм кустов, а шпалера заполненной, длина куста принята равной 200 см.
На ширину и высоту кроны куста влияют как система ведения куста, так и структура шпалеры. Высота куста меняется в пределах 144,9-172 см.
Самый высокий показатель по высоте куста отмечен при его формировании по типу полуукрывного веера КСХИ (172 см) при вертикальном ведении прироста и системе «Магарач-2» с двухъярусным размещением структурных элементов (166,5 см). Высота кустов остальных форм в пределах 150-160 см. По ширине кроны кусты на разных формах также отличаются в зависимости от способа ведения прироста и конструкции шпалеры. При вертикальном ведении прироста на полуукрывном веере КСХИ она равна 77,5 см, на кустах, сформированных по типу Омбрелла и длиннорукавная Каз. НИИПиВ- 1, несмотря на свободное ведение прироста, ширина кроны несколько ниже (96,2 и 104,1 см). Наличие только одной спаренной проволоки вызывает резко выраженное свободное свисание. Ширина кроны кустов на других формах колеблется в пределах 108,5- 126,4 см.
Таблица 1
Форма куста сорта Мерло | Средняя освещенность кроны кустов, тыс лк | |||||
Часы наблюдений | ||||||
7 | 10 | 13 | 16 | 19 | В среднем | |
Магарач-2 | 15,0 | 29,2 | 24,7 | 24,6 | 20,0 | 22,7 |
Магарач-Ильчер-1 | 15,7 | 29,0 | 22,4 | 27,4 | 17,4 | 22,4 |
Магарач-Ильчер-2 | 16,1 | 25,8 | 24,8 | 27,8 | 12,9 | 21,9 |
Таврическая полуукрывная | 16,3 | 36,7 | 30,6 | 32,5 | 17,5 | 26,7 |
Высокоштамбовый веер | 19,7 | 35,5 | 30,9 | 36,6 | 17,6 | 28,1 |
Омбрелла | 19,2 | 30,4 | 26,4 | 37,8 | 15,2 | 25,8 |
Длиннорукавная Каз. НИИПиВ-1 | 15,3 | 29,0 | 29,1 | 37,8 | 14,4 | 25.1 |
Полуукрывной веер КСХИ | 16,0 | 27,6 | 22,4 | 30,7 | 16,1 | 22,5 |
В результате самый большой объем кроны кустов наблюдался на Таврической высокоштамбовой полуукрывной форме (3,98 м3) и системе «Магарач-2» (3,78 м3), а самый низкий — на полуукрывном веере КСХИ (2,64 м3). Показатели объема кроны на остальных формах имеют близкие значения (3,1-3,5 м3).
Один из показателей, влияющий на работу листовой пластинки, — плотность расположения листьев в кроне куста. Так, плотность листового полога на полуукрывном веере КСХИ при вертикальном ведении прироста почти в 1,5 раза выше, чем при свободном (2,39 м2/м3). На кустах с Таврической высокоштамбовой полуукрывной формой и высокоштамбовым веером (120 см) плотность кроны несколько ниже по сравнению с другими формами, несмотря на большее число побегов (1,28 и 1,43 м2/), а на кустах с длиннорукавной формой Каз. НИИПиВ-1 и Омбрелла из-за наличия только одного яруса проволоки она выше (1,48 и 1,88 м2/м3), как и плотность листьев с верхней стороны кроны куста, что в конечном итоге будет сказываться на радиационном режиме.
Индексы листовой поверхности относительно горизонтальной (JLAJг) и вертикальной (JLAJв) проекции кроны отдельного растения имеют при вертикальном ведении прироста на полуукрывном веере КСХИ такие же высокие значения, как и плотность листового полога кроны (4,09 и 1,83 м2/м3). Довольно высоки значения этих показателей на кустах, сформированных по типу Омбрелла (3,02-1,81 м2/м3) и системе «Магарач-2» (2,85 и 1,96 м2/м3).
Радиационный режим виноградника.
Разные значения по плотности в кроне расположения листовой поверхности обусловили и различный характер освещенности кустов. По мнению А. Г. Сарнецкого (1981), недостаток прямого солнечного освещения усиливает процессы роста в ущерб плодоношению и снижает продуктивность фотосинтеза. По данным В. Бажова (1962), ориентированные с севера на юг ряды получают за день с восточной и западной стороны примерно одинаковое количество солнечной радиации: освещенность с солнечной стороны достигает 60 тыс. лк, на теневой стороне она значительно ниже (до 3-4 тыс. лк), а внутри кроны в полдень не превышает 400 лк.
Таблица 2
Согласно нашим исследованиям на радиационный режим виноградника большое влияние оказывают системы ведения и формирования кустов: на их освещенность определенным образом влияет каждая форма, имеющая свойственное ей пространственное размещение листовой поверхности и плотность расположения в кроне куста (табл. 1). Полученные данные показывают, что световой режим виноградника имеет два максимума (в 10 и 16 ч). В полдень наблюдается некоторое снижение освещенности кроны куста. В среднем за световой день освещенность кроны куста находится в пределах 22-28 тыс. лк. По световому режиму более благоприятные условия создаются в насаждениях с полуукрывными штамбовыми формами и длиннорукавным веером Каз. НИИПиВ-1 (25,1-18,1 тыс. лк), характеризующиеся более выраженным свободным свисанием побегов и меньшей плотностью расположения листьев в кроне куста.
Формирование кустов по системам «Магарач-2», «Магарач-Ильчер» с большой площадью листовой поверхности и плотностью листовой поверхности с односторонним размещением побегов имели несколько низкие значения по световому режиму (в пределах 22-23 тыс. лк). Близкие к этим способам ведения кустов значения отмечены на винограднике с полуукрывным веером КСХИ (22,5 тыс. лк). Различная степень освещенности кроны куста отразилась на продуктивности фотосинтеза.
Микроклимат виноградника.
По данным Т. Н. Турманидзе (1981), на температуру приземного горизонта воздуха, верхнего 20-сантиметрового слоя почвы и влажности воздуха в кроне куста основное влияние оказывает геометрическая структура самого виноградника.
В наших исследованиях разные формы кустов, имея различную структуру скелета куста, площадь листовой поверхности, режим освещенности в кроне куста, определенным образом повлияли на изменение микроклимата.
Наблюдения за температурой и относительной влажностью воздуха в кроне куста показывают некоторые различия в зависимости от его формы. Наиболее благоприятна для развития виноградной лозы относительная влажность воздуха в пределах 60-80%. Более низкая влажность ведет к непроизводительным потерям воды листьями и почвой, при этом нарушается естественное соотношение интенсивности физиологических процессов (А. Г. Сарнецкий, 1981). Относительная влажность воздуха в зоне размещения основной массы гроздей колеблется в пределах 54-64%. Самые низкие ее значения отмечены на высокоштамбовом веере (54%), а на остальных формах они близки к оптимальным.
Температура воздуха в кроне куста (зона размещения основной массы гроздей) в зависимости от формы куста меняется в пределах 25...28 °C. Высокая температура в кроне кустов, сформированных по системе «Магарач-Ильчер» с одним поводком (28,1 °C), объясняется близким размещением структурных элементов куста к поверхности почвы. На кустах с более выраженным свободным свисанием побегов зафиксированы меньшая листовая поверхность и более низкие относительная влажность и температура воздуха. Здесь также сказывается возможность свободного перемещения воздушных масс.
Меняется также температура верхнего 20-сантиметрового слоя почвы. В зависимости от формы куста почва в ряду и междурядье прогревается, имея максимум в полуденные часы. Сильно прогревается верхний 5-сантиметровый слой. В течение суток температура почвы в горизонте 15-20 см меняется незначительно и влияние формы куста незаметно.
Таким образом, микроклимат виноградника при различных системах ведения и формирования кустов имеет близкие значения, отрицательное влияние испытуемых форм на какие-либо показатели микроклимата не обнаружено: все они создают благоприятные условия для воздушно-светового питания растений.
Продуктивность фотосинтеза листового аппарата при различных способах ведения и формирования кустов.
Теоретической базой для повышения урожайности агрофитоценозов является комплексная теория фотосинтетической продуктивности (А. А. Ничипорович, 1963), рассматривающая условия оптимизации продуктивного процесса растений и возможности максимального использования поступающей лучистой энергии на формирование их урожая. Неотъемлемой частью такой методической основы при рассмотрении всех факторов, определяющих продуктивность растений, становится систематический подход, позволяющий применять анатомические методы и моделирование для изучения процессов формирования урожая растений.
Таким образом, цель наших исследований — определение характера взаимосвязей между поступающей ФАР и формированием фитосинтетического потенциала и урожая винограда в зависимости от системы ведения культуры. Как известно, максимальная продуктивность агроценоза возможна только при оптимальной его структуре и оптимальной площади листьев. Фотосинтетическая активность листьев зависит от количества поглощенной растениями энергии ФАР В свою очередь, на интенсивность поглощения ФАР влияют главным образом размеры листовой поверхности, ход ее формирования на протяжении вегетации и характер размещения листовой массы в пространстве. Как было выше отмечено, по радиационному режиму кусты различных форм находятся приблизительно в одинаковых условиях. При этом важную роль в повышении продуктивности насаждений играют площадь листовой поверхности, структура ее пространственного расположения, зависящая, в свою очередь, от системы ведения и формирования кустов.
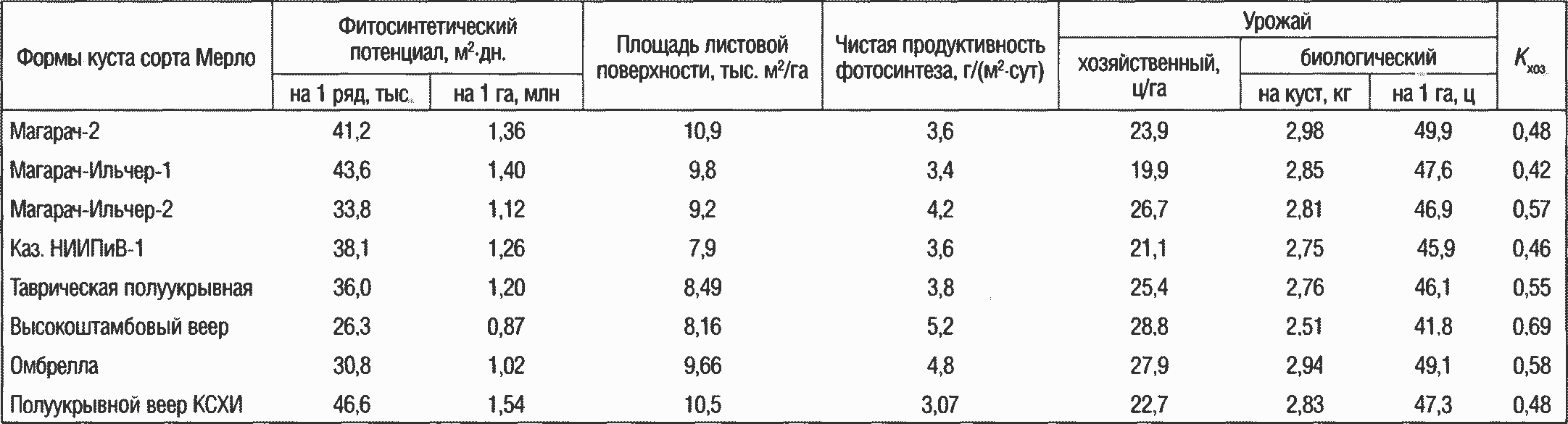
По данным А. Г. Амирджанова (1980) и Г. А. Сарнецкого (1981), площадь листовой поверхности в зависимости от системы ведения и формирования кустов составляет 8-43 тыс. м2/га. В наших исследованиях она колеблется в пределах 8,2-10,9 тыс. м2/га: самая высокая на кустах, сформированных по системе «Магарач-2» (10,9 тыс. м2/га) и по типу полуукрывного веера КСХИ (10,5 тыс. м2/га), на остальных формах — в пределах 8-9 тыс. м2/га.
Величина фотосинтетического потенциала на кустах с разной формой меняется так же, как и величина листовой поверхности, с увеличением которой при одинаковой продолжительности периода вегетации закономерно повышается и фотосинтетический потенциал: его самый высокий показатель зарегистрирован на полуукрывном веере КСХИ (1,54 млн м2-дн./га) и системе «Магарач-Ильчер» (1,40 млн м2-дн./га). На кустах с более выраженным свободным свисанием побегов фотосинтетический потенциал находится в пределах от 0,87 до 1,26 млн м2-дн/га (табл. 2).
Один из важных показателей фотосинтетической деятельности растений — чистая продуктивность фотосинтеза. По данным А. Г. Амирджанова (1980), она снижается на кустах с небольшой площадью листьев. Согласно нашим данным, площадь листьев сама по себе еще не определяет уровень чистой продуктивности фотосинтеза и зависит в основном от условий освещенности и нагрузки урожаем. Наблюдается тенденция к увеличению чистой продуктивности фотосинтеза с уменьшением фотосинтетического потенциала. Увеличение хозяйственной части
урожая приводит к повышению продуктивности фотосинтеза: ее самый высокий показатель отмечен на кустах, сформированных на высокоштамбовом веере и по типу Омбрелла [5,2 и 4,8 г/(м2-сут)], на полуукрывном веере КСХИ он равен 3,07 г/(м2-сут), а на остальных формах — 3,4-3,8 г/(м2-сут).
Коэффициент хозяйственной части урожая (Кхоз) характеризует долю сухой массы гроздей в общей массе годичной продукции растения: Кхоз = Ухоз/Убиол. Наибольшая величина Кхоз за вегетацию зафиксирована на штамбовых формах: от 0,55 ед. на Таврической
высокоштамбовой полуукрывной до 0,69 ед. на высокоштамбовом веере. Близкие к ним значения отмечены на полуукрывном веере КСХИ и системе «Магарач-2» (0,48 ед.).
На основании проведенных исследований сделан вывод, что по основным показателям фотосинтеза исследуемые формы кустов отличаются незначительно. В более благоприятных условиях находятся кусты с полуукрывной формой при свободном ведении прироста, а из укрывных форм выделяются кусты, сформированные по системе «Магарач-2».