СИНТЕЗ ОРГАНИЧЕСКОГО ВЕЩЕСТВА В ЛИСТЬЯХ ВИНОГРАДА И ЕГО ЗАВИСИМОСТЬ ОТ РАЗЛИЧНЫХ БИОЛОГИЧЕСКИХ И ЭКОЛОГИЧЕСКИХ ФАКТОРОВ
Накопление сахара в винограде зависит от работы листьев, их фотосинтетической активности и мероприятий, способствующих правильному распределению ассимилятов и их направлению к ягодам и другим органам растения. Окружающая виноград или воздействующая на него среда (экологические или агротехнические факторы) оказывает влияние на работоспособность и продуктивность листьев, что в итоге обусловливает его рост, определяет величину и качество урожая.
Листья винограда — практически единственный орган, способный синтезировать органические вещества из неорганических под воздействием солнечной энергии. Побеги и другие органы и ткани винограда, содержащие известное количество хлорофилла, также могут ассимилировать, но органическое вещество, синтезируемое ими, настолько незначительно, что практически не имеет значения для питания растений.
Суточные и сезонные изменения фотосинтеза. Все исследователи, изучавшие фотосинтез винограда, отмечают значительные изменения его интенсивности в течение суток и в различные фазы вегетации. Эти изменения обусловлены сложным влиянием ряда факторов внешнего и внутреннего характера.
Исследования Станеску (Stanescu, 1927, 1928, цит. по Georgescu, 1960) показали, что в суточном ходе фотосинтеза наблюдается несколько максимумов: первый — до полудня, второй — после полудня. Иногда к вечеру наступает и третий максимум.
А. С. Мержаниан (1951) также указывает на наличие двух максимумов. Другие авторы установили только один, максимум — около полудня, вследствие чего суточный ход фотосинтеза представляется как типичная одновершинная кривая (Бузин, 1951).
Хегедюш (Hegedus, 1952, 1956) изучал интенсивность ассимиляции в разное время дня и периоды года. По его данным, самая высокая ассимиляция установлена с 28 по 30 июня и с 12 по 14 июля и почти всегда в полуденные часы.
В. В. Васильева (1956) установила, что у одних сортов (Амурский и Альфа) максимум фотосинтеза наступает в дополуденные часы (7—11 ч), а у других (Буйтур и Коринка Мичурина) —в полуденные (11—15 ч), что сорта Амурский и Альфа в первую половину вегетации отличаются более высокой суточной продуктивностью фотосинтеза по сравнению с Буйтуром и Коринкой Мичурина. У первых не только интенсивнее накопление, но и энергичнее отток органического вещества.
По данным М. В. Моториной (1958), в условиях Московской области наиболее высокая интенсивность фотосинтеза у сортов Альфа и Амурский наблюдается между 8 и 10 ч, а у сорта Черный сладкий в полуденное и послеполуденное время. В общем у всех сортов понижение фотосинтеза после максимума незначительно.
Исследования М. В. Моториной показали, что суточный ход поглощения CO2, в значительной степени зависит от условий внешней среды. В дни с температурой 22—24° С и относительной влажностью воздуха не ниже 58—60% фотосинтез протекает по типу одновершинной кривой, интенсивность фотосинтеза находится почти на одинаковом уровне до 16 ч, после чего наблюдается ее снижение. Двухвершинная кривая характерна для дней с повышенной температурой (до 25— 26° С) и пониженной влажностью (до 40—43%) воздуха.
П. Г. Тавадзе (1961) также установил два максимума в суточном ходе фотосинтеза — первый между 9 и 10 ч, а второй между 16 и 17 ч со значительным понижением между 12 и 13 ч. Максимум фотосинтеза в дополуденные часы установлен и С. И. Кокиной (1937).
Результаты, полученные Георгеску (Georgescu, 1960), показывают, что в начальные фазы роста (до цветения), начиная с 7 ч утра, интенсивность фотосинтеза у сортов Алиготе, Мускат белый и Рислинг итальянский повышается к полуденным часам, достигая максимума в 13 ч. Во время цветения, как правило, максимальный фотосинтез наблюдается в 13, 16 и 19 ч. После цветения (18/VI), во время активного нарастания ягод (17—18/VII), во время созревания (10/IX) и после сбора урожая, как правило, устанавливаются два максимума — первый около 10 ч, а второй в 16 ч.

Рис. 57. Сезонные изменения продуктивности листьев:
7 — 4—6-й узел; 2 —8—12-й узел; 5 —15—18-й узел; 4 — 30—33-й узел
Автор отмечает некоторую зависимость между интенсивностью фотосинтеза и ходом суточных температур или влажностью воздуха. По его мнению, в начале вегетации максимумы фотосинтеза совпадают с более высокими температурами.
По данным Георгеску, фотосинтетическая активность листьев, расположенных в разных зонах по длине побега, неодинакова в отдельные фазы развития. До цветения (20/VI) наиболее активны листья, находящиеся между пятым и десятым узлом, затем примерно до начала созревания винограда (10—15/VIII)—листья 5—25-го узлов. Интенсивность фотосинтеза листьев, расположенных над 30-м узлом, выше, чем у основания побегов (1—5-й узел), но ниже, чем листьев средних зон побега.
К. Стоев, С. Добрева и Ю. Зейналов (1966) также изучали фотосинтетическую активность целых листьев в различных зонах по длине побега. Оказалось, что листья 4—6-го и 8—12-го узлов имеют почти одинаковый сезонный ход фотосинтеза: максимальная активность в конце июня—начале июля, а минимальная — в середине сентября. Листья 15—18-го и 30—33-го узлов проявляли высокую фотосинтетическую активность в августе и сентябре, из чего следует, что они играют существенную роль в питании растений и накоплении сахаров во второй половине вегетации (рис. 57).
Таким образом, на каждом этапе роста винограда существует определенная группа листьев, отличающаяся наиболее высокой фотосинтетической активностью и, очевидно, играющая наибольшую роль в питании растения.
Аналогичные исследования проведены А. А. Басанько и Е. Е. Горячевой (1964). Они установили, что у сорта Галан в июне наибольшей продуктивностью отличаются листья шестого яруса, в июле — девятого, в августе — пятнадцатого, а в октябре — тринадцатого.

Рис 58 Суточный ход фотосинтеза листьев разных ярусов 9 августа 1965 г. (А) и 20 сентября 1965 г. (Б):
1-1-6-й узел- 2-8-12-й узел; 3-15-18-й узел; 4-23-25-й узел; 5 - 30-33-й узел
К. Стоев, С. Добрева и Ю. Зейналов (1966), исследуя ассимиляционную активность листьев сорта Болгар, установили двухвершинный характер кривой суточного фотосинтеза (рис. 58). Характерно, что первый максимум фотосинтеза у более старых листьев наступает в ранние полуденные, а второй — в более поздние послеполуденные часы. У более молодых листьев первый максимум наступает ближе к полуденным, а второй — к более ранним послеполуденным часам. Это показывает, что в фотосинтетической активности листьев, расположенных в различных зонах основного побега, существует определенная координация.
К. Стоев, 3. Занков, И. Панайотов и Г. Найденова (1952) еще раньше установили, что координация ассимиляционной деятельности листьев сохраняет нормальный ритм в жизнедеятельности растений как в течение суток, так и в течение всего вегетационного периода, благодаря чему создаются наиболее благоприятные условия для оттока ассимилятов. Это положительно отражается на общей продуктивности растения.
Сезонные изменения фотосинтеза винограда изучали ряд авторов.
По данным В. В. Васильевой (1956), интенсивность фотосинтеза характеризуется общей тенденцией постепенного понижения к концу вегетационного периода, которую она объясняет старением листьев. Однако и в течение осенних месяцев интенсивность фотосинтеза остается сравнительно высокой. М. В. Моторина (1958) установила, что максимальной величины фотосинтез достигает в июне, после чего наступает постепенное понижение его интенсивности, хотя и во второй половине сентября он довольно высокий.
Тенденция к снижению интенсивности фотосинтеза во вторую половину вегетации установлена также К. Стоевым, С. Добревой и Ю. Зейналовым (1966).
Понижение интенсивности фотосинтеза во вторую половину вегетации отмечают А. М. Негруль и Л. Т. Никифорова (1958), Т. Г. Мазанашвили (1954), А.Н. Коленко и Н.П. Ерыгина (1953) и др.
В снижении интенсивности фотосинтеза листьев винограда во второй половине вегетации, вероятно, играют роль многие факторы. Среди них наибольшее значение, видимо, имеют изменения, наступающие в пластидах и хлорофилле по мере старения фотосинтетического аппарата. Известно, что старение листьев сопровождается деструкцией пластидов и уменьшением содержания хлорофилла, что, в конечном итоге, приводит к понижению интенсивности фотосинтеза. В связи с этим представляют интерес данные П. Г. Тавадзе (1952), показывающие значительное понижение содержания пигментов (хлорофилла, каротина, ксантофилла) в листьях после активного роста ягод. Вместе с этим значительно понижается и содержание воды в листьях, являющееся одним из важных условий фотосинтеза.
Предположение о зависимости между содержанием хлорофилла и интенсивностью фотосинтеза, однако, не подтверждается исследованиями Т. Г. Мазанашвили (1954) и М. Н. Чрелашвилк (1959).

Интенсивность фотосинтеза листьев, расположенных в различных зонах основного и бокового побегов. Один из внутренних факторов, влияющих на интенсивность фотосинтеза,— место расположения фотосинтезирующего органа и его возраст.
Одними из первых обратили на этот вопрос внимание Вилльштеттер и Штолл, 1918). Они показали, что молодые листья отличаются повышенной фотосинтетической активностью.
Определяя фотосинтез в листьях, расположенных у основания вызревшего побега, и в верхней, невызревшей части, И. Н. Кондо (1959) установил в конце вегетации (до листопада) значительно более высокую работоспособность молодых листьев. Ш. Ш. Чанишвили (1960) выявил наиболее высокую фотосинтетическую активность средних листьев (9,45 мг CO2 на 1 дм2/ч) по сравнению с верхушечными и низкорасположенными (соответственно 7,84 и 7,37 мг CO2 на 1 дм2/ч).
Л. Т. Никифорова (1959) разделила листья по длине побегов на три группы и изучала их фотосинтетическую активность, причем на отдельных этапах роста сохраняла на побеге одну группу листьев и удаляла другие. По ее данным, после цветения наиболее высокой работоспособностью отличаются нижние и средние листья, в период роста и созревания — средние, а затем верхушечные.
К. Стоев, 3. Занков, И. Панайотов, Г. Найденова (1952) установили закономерное перемещение наибольшей интенсивности фотосинтеза в течение вегетационного периода от листьев нижних и средних ярусов основных побегов к верхушечным и листьям пасынков и высказали предположение, что питание растения во второй половине лета и в конце вегетационного периода осуществляется листьями средних и верхних ярусов основного побега и более молодыми и жизнедеятельными листьями пасынков.
Позднее К. Стоев, С. Добрева и Ю. Зейналов (1966) показали, что в середине и к концу лета наблюдается резко выраженный акропетальный градиент в интенсивности фотосинтеза (рис. 59).
По данным С. Кондри (1962), листья верхних ярусов (15—30-й узлы) при замедлении роста побегов синтезировали лишь 44,1% сухого вещества, накопленного средними листьями, в то время как через 2—3 недели (при прекращении роста побегов) они синтезировали почти вдвое больше сухого вещества по сравнению со средними листьями. Осенью усиление фотосинтеза в верхних ярусах и молодых листьях выражено еще сильнее.
Одним из вопросов, вызывающих интерес у исследователей, является фотосинтетическая активность листьев боковых побегов и их роль в питании винограда.
В. Г. Александров, К. Ю. Абесадзе и Е. А. Макаревская (1929) считают, что пасынки при весьма слабой ассимиляции на единицу поверхности своих листьев испаряют почти столько же воды, сколько и листья нормальных побегов. Поэтому, по их мнению, пасынки являются своего рода полупаразитами лозы, непродуктивно расходующими ее водные запасы. Исходя из этого, авторы считают, что удаление пасынков, притом по возможности более раннее, желательно.
Исследования К. Д. Стоева и его сотрудников (1952), напротив, показали, что листья пасынков сорта Болгар обладают высокой интенсивностью фотосинтеза, которая особенно резко выражена во второй половине лета. В это время она значительно выше, чем у нижних и средних листьев. Пасынки, по мнению авторов, — полноценные органы винограда, выполняющие определенные функции, строго согласованные с функциями остальных частей растения и находящиеся в зависимости от всей его жизнедеятельности.
Сравнительно недавно опубликованы работы Ш. Ш. Чанишвили (1960, 1964), доказывающие методом меченых атомов углерода поступление в ягоды винограда ассимилятов, вырабатываемых листьями пасынков сорта Ркацители. По его данным, однако, из пасынков в ягоды поступает на много меньше ассимилятов (примерно в 5 раз), чем из листьев основного побега. Исходя из этого, Ш. Ш. Чанишвили считает пасынки неполноценными органами растения, которые целесообразно удалять.
Совершенно противоположные выводы делает М. Милосавлевич (1964). Применяя в своих исследованиях метод радиоактивных изотопов углерода (С14), он установил, что интенсивность фотосинтеза листьев пасынков значительно выше, чем листьев основного побега, и что необходимо отказаться от взгляда, согласно которому пасынки оказывают отрицательное влияние на развитие и урожайность винограда.
Н. Г. Дудник (1960) выявил, что листья пасынков подвоя Рипария ХРупестрис 101—14 еще в период активного роста (9/VI) существенно не отличались по интенсивности фотосинтеза от листьев основного побега. С прекращением роста (12/VIII) накопление углеводов в листьях пасынков протекало в 1,5 раза интенсивнее, чем в соответствующих по возрасту листьях основного побега. Не наблюдалось различий в интенсивности фотосинтеза наиболее молодых листьев основного побега и пасынков. Следует отметить, что в этих опытах рост пасынков был вызван искусственно. Листья естественно развивавшихся пасынков имели интенсивность фотосинтеза ниже, чем листья основного побега.
По мере старения листьев как основного побега, так и пасынка интенсивность фотосинтеза их снижалась.
В. Д. Корнейчук и Е. К. Плакида (1959) делают вывод, что пасынковые побеги способствуют накоплению пластических веществ в узлах основных побегов и увеличению числа плодовых почек. Они отмечают, что обязательное удаление одревесневших пасынков при обрезке и зеленых операциях в течение летнего периода безусловно неправильно.
Е. К. Плакида, А. Г. Мишуренко, В. И. Габович, Е. П. Нагорная и Л. Н. Сапожникова (1963) также подтверждают выводы о том, что пасынки являются полноценными органами. По их мнению, листья пасынковых побегов не уступают в своей работоспособности листьям основных побегов, а иногда и превышают их. Аналогичные выводы сделал также И. Н. Кондо (1960).
Учитывая разноречивость взглядов по этому вопросу, К. Д. Стоев и его сотрудники (1966) поставили цель еще раз проверить интенсивность фотосинтеза листьев пасынков, используя для этой цели инфракрасный газоанализатор, который дает возможность изучать динамику интенсивности фотосинтеза без удаления листьев с растения.
Таблица 75
Среднесуточная интенсивность фотосинтеза (в мг CO2 на 100 см2 листовой поверхности) листьев основного побега и пасынков (1964 г.)
|
|
| Разница | |
Дата анализа | Листья основного побега | Листья пасынков | в мг CO2 | в % фотосинтеза основного побега |
28/VII | 3,10 | 4,47 | 1,37 | 44,2 |
30/УП | 3,97 | 5,12 | 1,15 | 28,9 |
13/1Х | 2,14 | 3,87 | 1,73 | 80,3 |
20/1Х | 3,03 | 4,28 | 1,25 | 41,2 |
17/Х | 1,06 | 1,44 | 0,38 | 37,7 |
18/Х | 1,18 | 3,15 | 1,97 | 167,0 |
Результаты опытов показали, что интенсивность фотосинтеза листьев боковых побегов значительно выше, чем листьев основных побегов (рис. 60).
Среднесуточная интенсивность фотосинтеза листьев боковых побегов в течение трех месяцев была на 30—167% выше интенсивности листьев соответствующего яруса основных побегов (табл. 75).
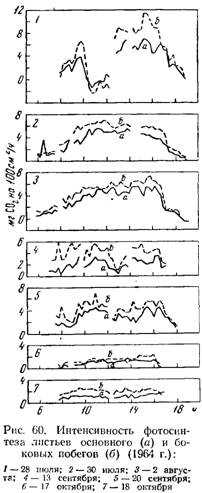
В большинстве случаев (60—95%) листья естественно развившихся пасынков у сорта Болгар обладают более высокой интенсивностью фотосинтеза, чем листья основного побега, находящиеся в основании соответствующего пасынка, причем разница бывает довольно значительной — 0,05—4,26 мг CO2 на 100 см2/ч. Отмечен ряд случаев, когда фотосинтетическая активность листьев на пасынках была такой же или немного ниже, чем листьев основного побега, однако это не дает основания считать пасынки полупаразитами.
Таким образом, можно считать доказанным, что листья боковых побегов (пасынков) играют значительную роль в питании винограда, ввиду чего их следует сохранять при его выращивании.
Причины более высокой фотосинтетической активности листьев боковых побегов не выяснены. Можно предположить, что одним из наиболее важных факторов является неодинаковое содержание пигментов (хлорофилла, каротина и ксантофилла) в листьях основных и боковых побегов, как и неодинаковая жизнедеятельность пластид в листьях различного возраста. В этом отношении представляют интерес исследования О. Т. Мишуренко (1963), установившего, что в начале вегетации листья боковых побегов содержат меньше пигментов. Позже, однако, содержание пигментов в этих листьях увеличивается на 50—60% по сравнению с листьями основных побегов.
Влияние некоторых экологических факторов и важнейших агротехнических мероприятий на интенсивность фотосинтеза.
Исследуя влияние температуры на фотосинтез, М. В. Моторина (1958) установила, что температурный оптимум зависит в значительной степени от сорта. У сорта Альфа фотосинтез повышается до температуры 20—25° С, после чего заметно падает (исследование проведено при освещенности более 20000 лк и относительной влажности воздуха 65— 75%). Понижение фотосинтеза у сорта Альфа с точки оптимума при падении температуры (с 24 до 14° С) происходит плавно и составляет около 3 мг CO2
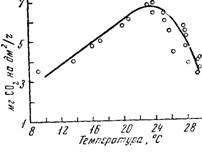
Рис. 61. Зависимость интенсивности фотосинтеза листьев от температуры
Аналогичная реакция на температуру наблюдалась и у сорта Амурский.
Установленный у сортов Альфа и Амурский оптимум не соответствует оптимальной температуре для других растений, выращиваемых в условиях средней географической широты. Эти два сорта отличаются высокой холодостойкостью и более коротким вегетационным периодом.
Сорт Черный сладкий отличается более длинным вегетационным периодом и значительно меньшей холодостойкостью. Как более теплолюбивый он отличается способностью переносить температуры выше и положительно реагирует на температуру 28—30° С. При высокой влажности воздуха температура 32° С все еще не оказывает на него угнетающего влияния.
Отмеченные различия, по мнению автора, соответствуют эколого-географическим условиям, в которых произошло формирование биологических качеств сортов.
Исследование Н. Селеджану (1962) показывает, что листья винограда, находящиеся в течение короткого времени (5 мин) в условиях высокой температуры (до 50°С), сохраняют свою способность поглощать CO2.
Значительное влияние на интенсивность фотосинтеза оказывает влажность воздуха. Исследования М. В. Моториной (1958) показывают, что понижение относительной влажности воздуха с 70 до 50% резко действует на интенсивность фотосинтеза. Специфичной реакции сортов к относительной влажности воздуха, однако, не установлено.
Исследования показали, что с усилением освещения до 20 000 лк, а для отдельных сортов и до 30 000 лк интенсивность фотосинтеза повышается (рис. 62). Дальнейшее усиление освещения не отражается на фотосинтетической активности листьев. К аналогичным выводам пришли и А. А. Басанько и Е. С. Горячева (1964).

Рис. 62. Зависимость интенсивности фотосинтеза сортов Черный сладкий (А) и Альфа (Б) от освещения:
1 — относительная влажность воздуха 65%. температура 22—26° С; 2 — относительная влажность 50—65%, температура 22—26° С; 3 — относительная влажность 50%, температура 26° С
Это показывает, что виноград, подобно другим растениям, в природных условиях использует свет не вполне продуктивно.
При постоянном освещении интенсивность фотосинтеза зависит от содержания влаги в воздухе. С увеличением относительной влажности интенсивность фотосинтеза повышается.
На фотосинтез влияет содержание золы в листьях. Ассимиляция CO2 понижается по мере снижения насыщения листьев водой (Bosian, 1964). Действие засухи, однако, смягчается подвоем. Например, понижение фотосинтеза у новой формы (Aeis), устойчивой к паразитическим грибам и филлоксере и выращиваемой на сухой почве (50% влагоемкости), составляет всего 0,92 мг CO2 на 100 см2/ч по сравнению с листьями сортов, выращиваемых на влажных участках (80% влагоемкости).
Фотопериод не оказывает непосредственного влияния на интенсивность фотосинтеза (Alleweldt, 1963).
Влияние других экологических факторов (географической широты, высоты над уровнем моря, состава почвы) на интенсивность фотосинтеза выяснено недостаточно. В литературе встречаются отдельные сведения о том, что местопроизрастание винограда оказывает известное влияние на интенсивность фотосинтеза (Чрелашвили, 1959).
Слабо изучено также действие на фотосинтез некоторых постоянных факторов при выращивании винограда: густоты посадки, подвоев, формировок и др. По мнению П. Г. Тавадзе (1953, 1961), ассимиляция наиболее высока при двусторонней шпалерной формировке, а В. И. Горбач (1958) считает, что при выращивании винограда на шпалерах с козырьком, в расстил, а также с восточной стороны при всех системах формирования.
Наиболее высокую интенсивность фотосинтеза П. Г. Тавадзе установил при густоте посадки 3300 кустов на 1 га. Как увеличение числа кустов до 8800—13 000 на 1 га, так и переход к формировкам типа Казенава приводит к понижению фотосинтеза. Это обстоятельство автор объясняет нарушением нормального соотношения между листовой поверхностью и корневой системой и ухудшением водного режима винограда.
А. Г. Мишуренко, В. И. Габович и Е. К. Плакида (1962) сообщают, что интенсивность фотосинтеза листьев сорта Серексия повышается при прививке на подвой 3309. При корнесобственной культуре или на подвое Рипария Глуар интенсивность фотосинтеза ниже.
У сорта Шасла белая интенсивность фотосинтеза повышается при прививке на подвой 101—14 по сравнению с подвоем 1616. У сорта Рислинг фотосинтез выше при прививке на подвой 3309 по сравнению с 1616, а у Саперави — на подвое 1309 по сравнению с подвоем 1202.
По мнению П. Т. Мамарова (1959), содержание хлорофилла в листьях в значительной степени повышается при прививке сортов Гымза и Мавруд на подвои Рупестрис дюЛо и Кобера 5ББ, по сравнению с прививкой на подвое Шасла хБерландиери 41 Б или же при корнесобственной культуре.
Сравнительно лучше изучено влияние некоторых агротехнических мероприятий (зеленых операций, орошения и удобрения) на фотосинтетическую активность листьев.
Наиболее высокую ассимиляцию С. И. Кокина (1937) установила при уменьшении числа листьев на 25% (прореживание) и удалении боковых побегов (пасынкование). Удаление только боковых побегов или боковых побегов и 50—65% листьев незначительно повышает интенсивность ассимиляции. Характерно, что уменьшение листовой поверхности сильно увеличивает отток веществ.
По данным С. И. Кокиной трудно определить, чем вызвано повышение фотосинтеза — стимуляцией его вследствие ранения или лучшим освещением. Понижение активности фотосинтеза при сильном уменьшении листовой поверхности она объясняет сильным высушиванием листьев вследствие действия света и ветра.
Л. Т. Никифорова (1959) также установила, что уменьшение числа листьев повышает интенсивность фотосинтеза, вследствие чего общее количество СО*, усвоенного одним побегом, уменьшается незначительно. По ее данным, уменьшение листьев до 10—11 на побег (примерно на 40% меньше, чем в контрольном варианте) повышает интенсивность фотосинтеза на 177—214%. Даже в варианте с оставлением 5 листьев над гроздью в некоторые фазы вегетации одним побегом усвоено наибольшее количество CO2. На основании этого автор приходит к выводу, что высокая интенсивность фотосинтеза, свойственная винограду, не используется полностью при обычных методах выращивания.
К. Д. Стоев и Д. Ц. Лилов (1956) изучали влияние прищипывания, пасынкования и чеканки основных и боковых побегов на различных этапах вегетации на интенсивность фотосинтеза. Результаты их исследования показывают, что применение зеленой обрезки приводит к временному повышению энергии фотосинтеза листьев винограда. Под влиянием прищипывания до цветения и во время цветения обнаруживается существенный сдвиг в сторону усиления фотосинтеза листьев, находящихся в зоне 4-го узла (до 145—244% от контроля). Значительное повышение энергии фотосинтеза (до 136—280%) отмечается и в вариантах, где прищипывание проводили в разгар цветения. В то же время фотосинтез листьев в зоне 8-го узла побега в тех же вариантах находился на уровне контрольных кустов, а в некоторых случаях был даже значительно ниже. Все это показывает, что прищипка верхушек побегов винограда до цветения и в разгар цветения благоприятно влияет на ход физиологических процессов только в зоне грозди.
Зеленые операции, применяемые позднее (ранняя чеканка), наоборот, усиливают фотосинтез расположенных выше листьев (в зоне 8-го узла до 180—225% от контрольных). В то же время в листьях зоны 4-го узла не отмечается усиление энергии фотосинтеза, за исключением случаев, где применялась сильная чеканка (на 12 листьев). Следует отметить, что влияние обрезки зеленых частей на фотосинтетическую активность листьев винограда, несмотря на некоторые отклонения, имеет хорошо выраженный временный характер.
Наибольший сдвиг в сторону усиления фотосинтеза листьев отмечается на 6—10-й день после проведения приема. На 15-й день интенсивность фотосинтеза под влиянием обрезки значительно ослабевает, хотя в некоторых случаях она все же выше, чем у листьев контрольных кустов.
Таким образом, действие обрезки зеленых частей на фотосинтез листьев винограда нестатично, а динамично.
С. Кондря (1962) также установил, что после чеканки интенсивность фотосинтеза заметно увеличивается. Это, по его мнению, зависит от сорта (у Совиньона больше, чем у Муската белого), интенсивности чеканки (при сильной чеканке больше, чем при слабой) и времени проведения операции. После чеканки интенсивность фотосинтеза наибольшая, затем она снижается и через 1 —1,5 месяца падает ниже контроля.
Известное влияние на интенсивность фотосинтеза оказывает также кольцевание. Исследования К. Д. Стоева и др. (1966) показывают, что кольцевание сорта Болгар подавляет фотосинтез листьев, причем это подавление имеет временный характер — депрессия длится до восстановления связи проводящей системы в месте кольцевания (рис. 63, 64).
Исследования С. А. Мельника и В. К. Анисимовой (1958) показывают, что листья плодоносных побегов фотосинтезируют сильнее, чем бесплодных. К концу вегетации, однако, фотосинтетическая активность листьев на бесплодных побегах в значительной степени повышается. На основании этого авторы считают, что при выращивании винограда необходимо обеспечивать определенное соотношение между плодоносными и бесплодными побегами.
X. Тодоров (1961) также установил, что листья плодоносных побегов фотосинтезируют интенсивнее по сравнению с листьями пасынков. Содержание хлорофилла в них ниже, а воды выше. Дыхание также интенсивнее у плодоносных побегов.
Как показали исследования А. А. Басанько, Е. С. Горячевой и Р. Т. Рябчуна (по сведениям Захаровой, 1964), значительное влияние на энергию фотосинтеза оказывает нагрузка побегами и гроздями.
Наличие грозди на побеге способствует известному повышению интенсивности фотосинтеза. Осуществляется ускоренный отток продуктов фотосинтеза в грозди, что ведет к стимуляции усвоения CO2. Чрезмерная нагрузка куста побегами и гроздями по мере увеличения листовой поверхности ведет к загущению и затенению части листьев и, следовательно, к ухудшению и относительному ослаблению ассимиляционной активности куста. Перегрузка куста листовой поверхностью, повышая затрату влаги и питательных веществ, поступающих из корней, может привести к ослаблению процессов фотосинтеза.
Влияние орошения на фотосинтез изучали немногие авторы. По данным И. Н. Кондо (1946), интенсивность фотосинтеза кустов при нормальном водоснабжении в утренние часы примерно в 2 раза, а в дневные часы — почти в 10 раз выше, чем кустов, выращиваемых в условиях плохого водоснабжения.
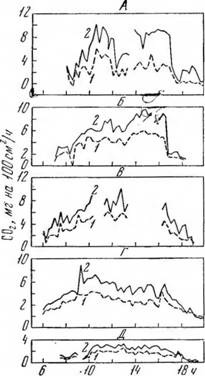
Рис. 63. Интенсивность фотосинтеза в зоне 5-го узла основного побега 1 июля (А), 7 июля (Б), 12 июля (В), 16 июля (Г) и 13 августа 1964 г. (Д):
I — кольцевание проведено 29 июня; 2 — без кольцевания
Весной, в начале лета, когда неорошаемые растения не испытывают еще недостатка влаги в почве, интенсивность фотосинтеза у них немного ниже, чем у орошаемых. Позже с ухудшением водоснабжения падает общий жизненный тонус богарных растений, ухудшается проветривание листьев, в несколько раз сокращается интенсивность фотосинтеза.
Исследования К. Стоева и Ю. Магрисо (1955) также показали, что орошение в значительной степени повышает интенсивность фотосинтеза. Наиболее значительное усиление фотосинтетической активности листьев отмечается на 3—8-й день после орошения.

Рис. 64. Динамика интенсивности фотосинтеза, листьев основного побега под влиянием кольцевания (а) и без кольцевания (б) по среднесуточным данным:
1 — листья 5-го узла; 2— листья 11-го узла; 3— листья 17-го узла

Рис. 65. Зависимость интенсивности фотосинтеза от величины сосущей силы и влажности почвы:
1 — сосущая сила орошаемых кустов (в атм.); 2 — сосущая сила неорошаемых кустов; 3 — фотосинтез листьев орошаемых кустов (в г сухого вещества); 4 — фотосинтез листьев неорошаемых кустов; 5 — влажность почвы орошаемого участка; 6 — влажность почвы неорошаемого участка
В некоторых случаях авторы наблюдали повышение фотосинтеза в 2—10 раз по сравнению с фотосинтезом листьев на неорошаемых участках.
К. Стоев и Ю. Магрисо установили также, что существует положительная корреляция между интенсивностью фотосинтеза, с одной стороны, и сосущей силой кустов и влажностью почвы, с другой.
Чем ниже сосущая сила и выше влажность почвы, тем выше интенсивность фотосинтеза. Орошаемые кусты всегда имеют сосущую силу ниже, а энергию фотосинтеза выше (рис. 65). Иногда за незначительными изменениями влажности почвы следуют заметные изменения сосущей силы, что сказывается на интенсивности фотосинтеза.
Сосущая сила как показатель, характеризующий до некоторой степени энергетическое состояние воды, выражает физиологическую активность воды в клетках паренхимы орошаемых и неорошаемых кустов. Более высокое содержание воды в почве оказывает положительное влияние на фотосинтез путем изменения осмотических свойств клеток ассимиляционной ткани до наступления ощутимых изменений в оводненности листьев.
Данные о влиянии удобрения (корневого и внекорневого) на фотосинтез имеют противоречивый характер. Не вполне выяснено также влияние инсектицидов и фунгицидов на фотосинтетическую активность листьев.