Изменение фотосинтеза листьев, расположенных на различных ярусах основного и бокового побегов1
Одним из внутренних факторов, оказывающих сильное влияние на интенсивность фотосинтеза, является местоположение фотосинтезирующего органа, включая его возраст. По этому вопросу в литературе встречаются весьма скудные, отрывочные данные.
Насколько нам известно, первыми исследователями, установившими, что молодые листья отличаются повышенной фотосинтетической активностью, являются Willstatter и Stoll (1918). Определяя фотосинтез в листьях, расположенных у основания вызревшего побега и в верхней, невызревшей части, К о н д о (1959) установил в конце вегетации (до листопада) значительно большую работоспособность более молодых листьев. Чанишвили (1960) устанавливает, что наиболее высокой активностью фотосинтеза отличаются средние листья (9,45 mg CO2/dm2 на час), в то время как верхушечные и низко расположенные листья ассимилируют слабее — соответственно 7,84 и 7,38 mg CO2/dm2 на час.
Никифорова (1959) разделяет листья по длине побега на три группы и изучает их фотосинтетическую активность, причем на отдельных этапах роста сохраняет на побеге одну группу листьев и удаляет другие листья. Она устанавливает, что после цветения наиболее высокой работоспособностью отличаются нижние и средние листья, в период роста и созревания — средние, а затем верхушечные.
Довольно полная разработка вопроса о метамерных изменениях фотосинтеза листьев виноградной лозы проведена в исследованиях К. Стоева с сотрудниками. В одной из наиболее ранних работ (Стоев и др., 1952) они установили, что в течение вегетационного периода интенсивность фотосинтеза закономерно перемещается от основных к верхушечным листьям и боковым побегам, и высказали предположение, что питание растения во второй половине лета и в конце вегетационного периода осуществляется листьями средних и верхних ярусов основного побега и листьями боковых побегов (пасынков), более молодых и жизнедеятельных.
В более поздних исследованиях Стоева с сотр. (1966а) вопрос о метамерных изменениях фотосинтеза получил дальнейшее развитие. Не останавливаясь в подробностях на полученных результатах, упомянем здесь только данные рис. 4, где представлена интенсивность фотосинтеза листьев сорта Болгар пяти ярусов побега (I — 4—6-й узел; II — 8—12-й узел; III —15—18-й узел; IV — 23—25-й узел; V — 30—33-й узел), определенная в два срока — 9 августа и 20 сентября 1965 г. Эти данные показывают, что в середине и к концу лета наблюдается резко выраженный акропетальный градиент в интенсивности фотосинтеза.
Статистическая обработка показывает, что колебания значений интенсивности фотосинтеза довольно велики, вследствие чего различие между двумя соседними ярусами через пять листьев не всегда ясно доказано. Различие в интенсивности фотосинтеза, однако, реально и полностью доказуемо для листьев через каждые 10—12 узлов побега.
Kriedemann (1968) подтверждает, что активность фотосинтеза выше у более молодых листьев по сравнению с более старыми.
1 Материал разработан К. Д. Стоевым и С. Добревой.
В то же время он устанавливает, что у наиболее молодых, еще не вполне сформировавшихся листьев (верхушечных) фотосинтез понижен.
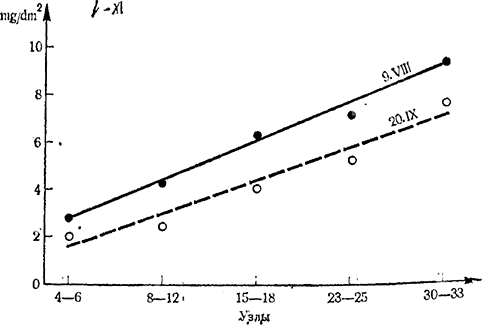
Рис. 4. Метамерные изменения интенсивности фотосинтеза (mg СO2 /dm2 на час) листьев сорта Болгар по длине побегов
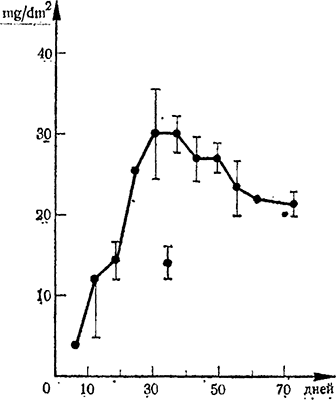
Рис. 5. Взаимосвязь между возрастом листьев (в днях) и интенсивностью фотосинтеза (mg COa/dm2 на час)
Кондря (1962) установил, что листья верхних ярусов (15—30-й узел) при замедлении роста побегов синтезировали лишь 44,1% сухого вещества, накопленного в средних листьях, в то время как через 2—3 недели (при прекращении роста побегов) они фотосинтезировали почти вдвое больше (86,9— 186,9%) по сравнению со средними листьями. Осенью усиление фотосинтеза в верхних ярусах и молодых листьях еще сильнее выражено.
Позже Кriеdemann с сотр. (1970) показали, что максимальная фотосинтетическая активность листьев обнаруживается в возрасте 40 дней, после чего снижается (рис. 5). По мнению авторов повышение фотосинтеза листьев в процессе их развития не обусловлено (не связано атрибутивно) увеличением концентрации хлорофилла, спектральными параметрами или же какими-либо значительными анатомическими изменениями.
Одним из вопросов, вызывающих повышенный интерес среди исследователей, является вопрос о фотосинтетической активности листьев боковых побегов и их роль в питании виноградной лозы.
Александров с сотр. (1929) считают, что пасынки при весьма слабой ассимиляции на единицу поверхности своих листьев испаряют почти столько же воды, сколько и листья нормальных побегов. Поэтому по их мнению пасынки являются своего рода полупаразитами лозы, непродуктивно расходующими ее водные запасы. Исходя из этого, авторы считают желательным удаление пасынков, притом по возможности более раннее.
Исследования Стоева с сотр. (1952), наоборот, доказали, что листья пасынков сорта Болгар обладают высокой интенсивностью фотосинтеза. Повышенная интенсивность фотосинтеза пасынков особенно резко выражена во второй половине лета, когда она значительно выше, чем у нижних и средних листьев.
Пасынки, по мнению авторов, являются полноценными органами виноградной лозы, выполняющими определенные функции, строго согласованные с функциями остальных частей растения и находящиеся в зависимости от всей его жизнедеятельности.
Чанишвили (1960, 1964) снова возвращается к этому вопросу и, применяя методы меченых атомов углерода, утверждает, что радиоактивные ассимиляты сорта Ркацители поступают в ягоды из пасынков в намного меньшей степени (примерно в 5 раз), чем из листьев основного побега. Исходя из этого, Чанишвили заключает, что пасынки являются неполноценными органами виноградного растения и что их целесообразно удалять.
Совершенно противоположные выводы делает Milosavlievic (1964). Применяя в своих исследованиях тот же метод радиоактивных изотопов углерода (14С), он устанавливает, что интенсивность фотосинтеза листьев пасынков значительно выше, чем листьев основного побега. В заключение автор отмечает, что необходимо отказаться от взгляда, согласно которому пасынки оказывают отрицательное влияние на развитие и урожайность лозы.
Дудник (1960) установил, что листья пасынков подвоя Рипария х Рупестрис 101-14 еще в период активного роста (9/VI) существенно не отличались по энергии фотосинтеза от листьев основного побега. С прекращением роста (12/VIII) углеводы в листьях пасынков накоплялись в 1,5 раза интенсивнее, чем в соответствующих по возрасту листьях основного побега. Не наблюдалось различий в интенсивности фотосинтеза наиболее молодых листьев основного побега и пасынков. Следует отметить, однако, что эти проявления листьев боковых побегов наблюдались при условии искусственно вызванного роста пасынков. Листья самовольно развившихся пасынков обладали более низкой энергией фотосинтеза, чем листья основного побега. По мере старения снижалась интенсивность фотосинтеза как листьев основного побега, так и листьев пасынка.
Корнейчук и Плакида (1959) делают вывод, что пасынковые побеги способствуют накоплению пластических веществ в узлах основных побегов и увеличению числа плодовых почек из соцветия. В заключение авторы отмечают, что обязательное удаление пасынков при обрезке и зеленых операциях в течение летнего периода безусловно неправильно.
Плакида с сотр. (1963) также подтверждают выводы о том, что пасынки являются полноценными органами виноградной лозы. По их мнению, листья пасынковых побегов не уступают в своей работоспособности листьям основных побегов, а иногда и превосходят их. К аналогичным выводам приходят также Кондо (1960) и Сапожникова (1963).
Учитывая разноречивость взглядов по этому вопросу, Стоев с сотр. (1966) поставили себе целью еще раз проверить интенсивность фотосинтеза листьев пасынков, используя для этой цели инфракрасный газоанализатор, позволяющий изучать динамику интенсивности фотосинтеза без удаления листьев с растения. Часть полученных результатов представлена на рис. 6. Эти результаты показывают, что интенсивность фотосинтеза листьев боковых побегов сорта Болгар значительно выше, чем листьев основных побегов.
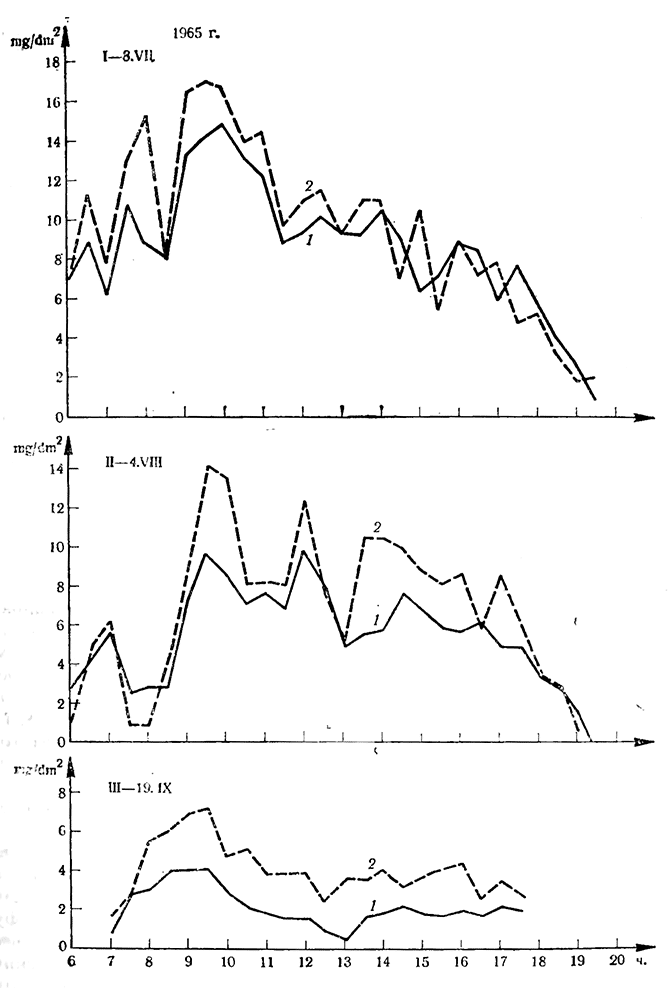
Аналогичные результаты получены и для сорта Ркацители. Из графика, построенного по средним арифметическим из шести определений (рис. 7), видно, что интенсивность фотосинтеза листьев пасынков близка к интенсивности фотосинтеза листьев основных побегов. Однако в повторностях наблюдались известные колебания. В течение трех месяцев интенсивность фотосинтеза листьев боковых побегов была на 30—167% выше интенсивности листьев соответствующего яруса основных побегов.
Таблица 1
Сопоставление интенсивности фотосинтеза листьев пасынков и основных побегов
1965 | Ярус листьев основного побега | Вероятность встречаемости более высокой интенсивности фотосинтеза листьев пасынков, | Уровень | Разница интенсивности фотосинтеза, mg CO2/dm2 на час |
7/VII | 4— 6-й узел | 90 | 0,10 | 0,09—2,81 |
3/VI1I | 4— 6-й узел | 95 | 0,05 | 0,19—4,41 |
18/IX | 4— 6-й узел | 80 | 0,20 | 0,05—1,82 |
8/VII | 10—12-й узел | 60 | 0,40 | 0,27—2,87 |
4/VIII | 10—12-й узел | 60 | 0,40 | 0,38—2,72 |
19/IX | 10—12-й узел | 90 | 0,10 | 0,36—4,26 |
В большинстве случаев (60—95 %) листья самовольно развивавшихся пасынков у сорта Болгар обладают более высокой интенсивностью фотосинтеза, чем листья основного побега, находящегося у основания соответствующего пасынка, причем разница бывает довольно значительной — 0,05—4,26 mg CO2/dm2 на час (табл. 1). Действительно, в ряде случаев фотосинтетическая активность листьев пасынков была такой же или немного ниже, чем листьев основного побега, однако это ни в коем случае не дает основания считать их полупаразитами.
Панделиев, с сотр. (1977) также установили, что интенсивность фотосинтеза листьев пасынков значительно выше, чем листьев основного побега.
На основании проведенных исследований можно считать, что листья боковых побегов (пасынков) играют значительную роль в питании виноградной лозы, ввиду чего их следует сохранять при ее выращивании.
Причины повышенной фотосинтетической активности листьев боковых побегов не выяснены. Можно предположить, что одним из наиболее важных факторов является неодинаковое содержание пигментов (хлорофилла, каротина и ксантофилла) в листьях основных и боковых побегов, как и неодинаковая жизнедеятельность пластидов в листьях различного возраста.
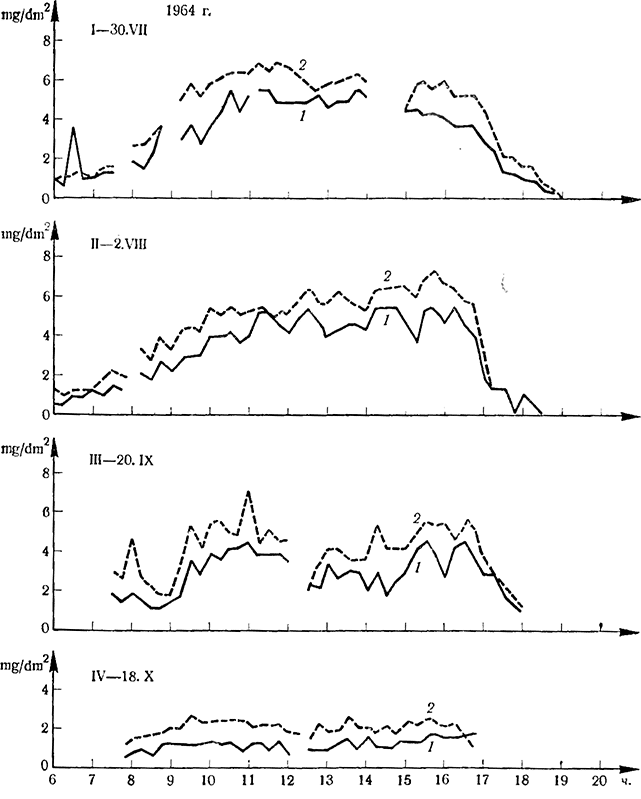
Рис. 6. Интенсивность фотосинтеза листьев (mg CO2/dm2 на час) основного побега и пасынка (1964 и 1965 гг.)
1964 г.: I — листья X яруса основного побега (7) и листья V—VIII яруса соответствующих пасынков (2); II — листья пасынков (2); III, IV — листья IV—V и X—ХII яруса основного побега (7) и листья III—VI ., I, II, III — средняя зона побега (10—12-й узел); IV—V яруса основного побега (I) и листья средней части соответствующих; 1 — листья основного побега узла соответствующих пасынков (2), 2 — листья пасынков
В этом отношении представляют интерес исследования Мишуренко (1963), установившего, что в начале вегетации листья боковых побегов содержат меньше пигментов. Впоследствии, однако, содержание пигментов в этих листьях увеличивается на 50—60% по сравнению с листьями основных побегов.

Рис. 7. Интенсивность фотосинтеза листьев сорта Ркацители (mgCO2/dm2 на час)
1 — основной побег; 2 — пасынок
Что касается разноречивых данных об интенсивности фотосинтеза листьев основного побега и пасынка сорта Ркацители, как нам представляется, они вызваны тем, что способность к развитию боковых побегов у этого сорта весьма невелика, причем она сильно зависит от условий выращивания. Не исключено, что на бедных и сухих почвах Грузии листья слаборазвитых пасынков сорта Ркацители ведут себя иначе, чем у других сортов.