Наиболее подробные исследования в связи с выяснением вопросов, касающихся передвижения ассимилятов, в прошлом проведены Hale и Weaver (1962) и Чанишвили (1964).
Путем подкормки 14СO2 листьев различных ярусов побега в различные этапы вегетации и последующего изучения передвижения ассимилятов методом радиоавтографии отдельных частей и органов виноградной лозы Hale и Wеavеr (1962) получили довольно полное представление о восходящем и нисходящем токе ассимилятов. В обобщенном виде их выводы представлены схематически на рис. 16.
По мнению авторов молодой лист ведет паразитическое существование и использует готовые продукты питания. Leonard и Weaver (1961) установили, что до достижения половины своей нормальной величины лист не в состоянии выделять ассимиляты для использования другими частями виноградной лозы.
Как показывает рис. 16 (а), движение ассимилятов в начале акропетальное. После формирования новых, молодых листьев, принимающих функцию акропетального отправления ассимилятов, более старые и почти нормально развитые листья отправляют ассимиляты в базипетальном направлении вниз к грозди. Это распределение ассимилятов установлено авторами в период цветения и до цветения виноградной лозы (б). К этому времени имеется 10—12 листьев, отправляющих ассимиляты в базипетальном направлении.
После опыления цветков ассимиляты, выработанные листьями под гроздью, расходятся в двух направлениях — вниз к основанию куста и вверх к грозди. Листья, расположенные над гроздью, также отправляют ассимиляты в двух направлениях — к грозди или верхушке побега, в зависимости от их местоположения и степени развития. Движение ассимилятов из листьев в секторе 8—9-го узла полностью базипетальное, а в секторе 12—13-го узла — главным образом акропетальное.
В начале созревания винограда (рост побегов в длину приостанавливается) ассимиляты из всех листьев, включительно и расположенных в 10 cm от верхушки побега, направляются вниз к грозди (в). Такая же картина наблюдается и позже (во время уборки урожая).
Таким образом, движение ассимилятов, выработанных в листьях, происходит в двух направлениях — вверх к конусу нарастания и вниз к винограду и к основанию куста.

Рис. 16. Схематическое изображение передвижения ассимилятов вверх и вниз по побегу
До цветения ассимиляты направлены главным образом к конусу нарастания и молодым листьям—кисти слабые потребители ассимилятов. После опыления, однако, главным потребителем становятся растущие ягоды, и к ним направляются ассимиляты из основной части листьев, расположенных под и над ними. Конус нарастания снабжают ассимилятами 7—8 верхушечных листьев, из которых только несколько нормально развиты.
Впоследствии Jaquinet и Simon (1977) представили передвижение ассимилятов вверх и вниз по побегу, как это показано на рис. 17. Из схемы видно, что по мере ослабления роста побегов непрерывно уменьшается число листьев на питание и нарастание верхушки побегов. Вместе с этим в процессе формирования винограда и созревания ягод закономерно увеличивается число листьев, отправляющих продукты фотосинтеза вниз, которые растение утилизирует на накопление сахара в винограде, на вызревание побегов и запасание углеводами в корневой системе и многолетних частях.
Hale и Weaver (1962) установили некоторые особенности и в движении ассимилятов к пазушным почкам, верхушкам боковых побегов, усикам, камбию и другим органам, тканям и частям виноградной лозы.

Рис. 17. Схематическое изображение передвижения ассимилятов вверх и вниз по побегу в процессе его роста и созревания ягод
1 — начало роста побегов; 2 — до цветения; 5 — после цветения; 4 — рост ягод; 5 — начало созревания; 6 — зрелость ягод
Они считают их второстепенными потребителями, которые не оказывают влияния на движение ассимилятов в продольном направлении. Следует отметить, что движение ассимилятов к второстепенным потребителям происходит со стороны обработанного листа, если только главные потребители не изменяют их направления. Это авторы называют "линейностью переноса".
Явление линейности в переносе ассимилятов установили также Стоев с сотр. (1974). По их данным "линейность" настолько сильно выражена, что путь ассимилятов отмечается радиоактивными продуктами 14СO2 преимущественно с той стороны двухлетнего побега, многолетних частей и корней, на которой по ортостихному расположению находился побег текущего прироста, обработанный меченым углеродом.
В фазе начала созревания в линейности переноса ассимилятов наступают изменения и отмечается интенсивный обмен продуктов фотосинтеза между разными побегами. Несколько позже, во время созревания ягод винограда, линейность в передвижении ассимилятов вновь восстанавливается.
Линейность переноса веществ установлена и в восходящем направлении. Обрабатывая корни винограда 14С сахарозой, радиоактивным бикарбонатом натрия и 32Р, всегда обнаруживали линейность переноса веществ. Явление это было настолько ярко выражено, что заранее можно было бы предсказать, в каком месте надземной части появится радиоактивный продукт, введенный в корни.
Лебедев (1973) посредством инфильтрации фуксина и метиленового синего в корни и стеблевые части также подтверждает существование „линейности“ в передвижении веществ. Далее Иванчев и Стоев (1978) снова установили, что ассимиляты побега, обработанного 14СO2, в большинстве случаев поступают в один из основных корней, находящихся с той стороны стрелки, на которой находится подкормленный побег. В тех случаях, когда 14СO2 обрабатываются два побега, находящихся на противоположных рукавах куста, радиоактивные ассимиляты идут вниз к корневой системе по двум нисходящим токам и поступают в корни, расположенные с соответствующих сторон меченых побегов.
Иногда путь ассимилятов в зависимости от сложности структуры куста проходит двумя, тремя и более токами, которые проникают в отдаленные друг от друга корни. При этом перераспределение ассимилятов осуществляется в зоне головки куста. Перераспределение может совершиться и в нижней части подземного стебля. В таком случае ассимиляты иногда поступают в корни, находящиеся в противоположной стороне от обработанного радиоактивным углеродом побега.
Продолжительность выноса основной части меченых ассимилятов из листьев — около 48 ч. Время оттока несколько дольше, если ассимиляты поступают из верхних листьев, и короче для листьев базальной части побегов. Установлено также, что листья расположенных выше гроздей направляют ассимиляты преимущественно в верхнюю гроздь, листья же в зонах гроздей отправляют ассимиляты к нижней грозди. Я годы, над которыми нет листьев, в фазе начала созревания потребляют ассимиляты из соседних побегов.
Большое значение имеет установленный Stoev и Ivantchev (1977) факт, что в период начала созревания винограда отток ассимилятов из кустов с нормальной нагрузкой к корневой системе и многолетним ветвям временно приостанавливается и все продукты фотосинтеза направляются к созревающим ягодам. На рис. 18—21 представлено графически передвижение и транслокация радиоактивных ассимилятов до начала созревания (2/VII) и в фазе начала созревания ягод (29/VIII). Как видно, интенсивное накопление радиоактивных ассимилятов в ягодах и передвижение и транслокация меченых продуктов фотосинтеза происходят в фазе начала созревания ягод. Результаты этих исследований, как нам представляется, являются вкладом в изучение проблемы, долгое время волновавшей исследователей — почему в фазе начала созревания ягод наступает интенсивное ("резкое", "скачкообразное") накопление сахаров в ягодах винограда и каков источник углеводов для синтеза сахаров в ягодах. В кустах с низкой нагрузкой отмечалось некоторое поступление ассимилятов и к корневой системе.
Исследованиями Stoev и Ivantchev (1977) установлено также начало восходящего тока углеводов весной из корневой системы. В начале марта уже начинается перемещение радиоактивных продуктов из корневой системы к надземным органам. Этот факт дал основание авторам считать, что биологическим началом вегетации является гидролиз крахмала в корнях и передвижение продуктов его распада и что это происходит значительно раньше наступления сокодвижения.
По мнению Соунсона ассимиляты передвигаются со скоростью 1 cm/min. Чанишвили устанавливает, что в нисходящем направлении ассимиляты передвигаются со скоростью 34—44cm/h, а в восходящем — соскоростью 15—17cm/h. Передвижение ассимилятов на значительные расстояния (15 футов за два дня) из подкармливаемого листа установили также Meynhard и Мakаn (1963).
Вклад в изучение проблем передвижения ассимилятов вносят Коb1еt, Perret (1971, 1972), Коb1еt (1975, 1977), причем их выводы направлены преимущественно на решение вопросов, имеющих значение для практического виноградарства.
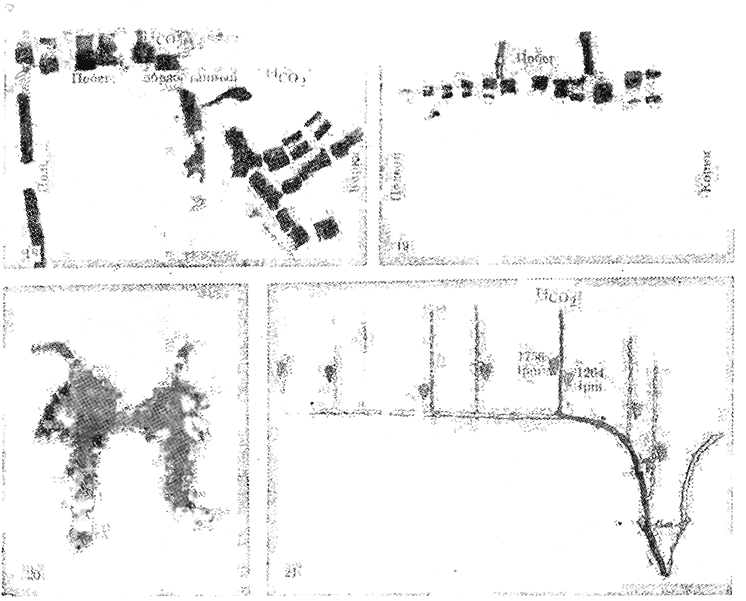
Рис. 18. Авторадиография передвижения ассимилятов до начала созревания ягод винограда (2/VII 1970). Сверху побег, обработанный 14СO2; слева радиоактивные ассимиляты в подземном стебле; справа транслокация ассимилятов в корнях линеарно со стороны побега, обработанного 14СO2
Рис. 19. Транслокация радиоактивных продуктов фотосинтеза в фазе начала созревания ягод (29/VIII 1970)
Рис. 20. Передвижение радиоактивных ассимилятов не обнаружено в подземном стебле и в корнях винограда — они локализированы лишь в побегах и гроздях
Рис. 21. Транслокация радиоактивных продуктов ассимиляции после фазы "начало созревания"; передвижение возобновлено вниз к корням
В работе, опубликованной в 1971 г., Koblet и Perret сообщают результаты серии опытов с радиоактивным углеродом 14СO2 по изучению роли пасынков в питании винограда. Сущность опытов заключается в следующем. На 21 кусте автор сохранил пасынки, а на 17 проводил пасынкование. Часть пасынков не имела соцветий или же авторы их удаляли, другие пасынки оставлялись с соцветиями. Экспозиция листьев пасынка в атмосфере, насыщенной 14СO2, длилась 60 min; спустя 24 h производили радиоавтографию основных побегов.
Результаты показали, что листья пасынков без соцветий, соответственно без гроздей, выносят ассимиляты к гроздям основных побегов (рис. 22), в то время как пасынки с гроздями не всегда направляют ассимиляты к основным побегам (рис. 23).

Рис. 22. Передвижение ассимилятов из пасынков без гроздей
а — неплодоносящий (бесплодный) пасынок; б — виноград с пасынками, снятый за 6 дней до опыта

Рис. 23. Передвижение ассимилятов из пасынков с гроздями
а — второй лист обработан 14СO2; б — четвертый лист обработан 14СO2
Это показывает, что соцветие пасынка является основным потребителем ассимилятов. Удаление соцветий (грозди) не изменяет свойств плодоносящего пасынка, он сохраняет ассимиляты для себя.
Установлено также, что передвижение ассимилятов из пасынка происходит линеарно к гроздям основных побегов. При этом направление ассимилятов не меняется, если лист в зоне грозди снят. Существенное значение имеет тот факт, что виноград кустов с пасынками отличался большей сахаристостью на 3,6° по Эксле. На этом основании автор делает вывод о значительной роли пасынков в повышении качества винограда.

Рис. 24. Экспорт ассимилятов из побегов двойников
К сожалению, Koblet остались неизвестными исследования, проводившиеся в 1952 и 1966 гг. Стоевым с сотрудниками, которые выявили высокую фотосинтетическую активность листьев пасынков и утвердили полноценность побегов пасынков в питании винограда, поэтому он ссылается на более поздние работы (Hale, Weaver, 1962; Sсhoff1ing, 1967; Koblet, 1969), которые лишь подтверждают ранее сделанные выводы. В обзоре настоящей работы аннотированны исследования Корнейчука и Плакиды (1959), Кондо (1960), Дудника (I960), Сапожниковой (1963), Milosavlievic (1964), подтвердившие несколько позже на других объектах и в других районах, что интенсивность фотосинтеза листьев пасынков такая же, как интенсивность фотосинтеза листьев основных побегов, а иногда и выше.
Интерес вызывает исследование Koblet и Perret (1972), изучавшие передвижение ассимилятов в двух побегах, выросших из одного глазка, из побегов, не подвергавшихся прищипыванию и прищипнутых, а также из жирующих побегов.
Результаты наблюдения после обработки побегов двойников радиоактивным углеродом (14СO2) представлены на рис. 24. Как видно из левой части графика, при обработке 14СO2 плодоносящего побега двойника установлен слабый отток в смежные побеги. При обработке неплодоносящего побега двойника (правая часть графика) обнаруживается значительный отток в побег двойника. Иногда авторы наблюдали, что отток ассимилятов из плодоносящего побега двойника ограничивается лишь соседним побегом. Наоборот, иногда отток из неплодоносящего побега двойника направляется к соседним побегам. Если обработка проводится в фазе начала созревания винограда, передвижение в соседние побеги еле заметно, что объясняется интенсивным потреблением ассимилятов ягодами винограда.

Рис. 25. Передвижение ассимилятов под влиянием прищипывания верхушки побегов (12/VII 1971)
а — без прищипывания; б — с прищипыванием
Стоев и Иванчев (1977) изучали взаимосвязи между плодоносящими и бесплодными побегами в питании виноградной лозы. Они установили, что ассимиляты плодоносящих побегов идут главным образом на питание гроздей и ягод, и эти побеги проявляют сильно выраженную "автономность". Ассимиляты неплодоносящих побегов используются растением преимущественно на вегетативный рост и накопление в корневой системе запасных углеводов. В фазе начала созревания ягод, однако, они отдают почти все ассимиляты на накопление сахаров в ягодах плодоносящих побегов.
Очень интересные результаты получили Koblet и Perret (1972) при прищипывании верхушки побегов. Как видно из данных рис. 25, прищипывание побега вызывает интенсивный отток ассимилятов не только к соцветиям этого же побега, но и к соседним побегам плодовой стрелки.
Эти сведения показывают, что прищипывание верхушки побегов является надежным средством перераспределения ассимилятов и направления их к боковым точкам роста, прежде всего, к соцветиям. Эта операция, как известно из прежних исследований, является эффективным мероприятием против осыпания цветков и завязей винограда, поскольку направляет временно, до возобновления роста в длину, все ассимиляты к соцветиям. (Стоев, 1944— 1945; Мержаниан, 1951 и др.)
Обнаружен также отток ассимилятов и у неподвергнутого прищипыванию побега, однако он ограничивается главным образом соцветиями того же побега и незначителен в соседние побеги. В одном из опытов отток к соседним побегам вообще не обнаружен. В случае, что в соседстве с неприщипнутым побегом имеются слабо развитые плодоносящие побеги, часть ассимилятов направляется к ним.
Жирующие побеги направляют выработанные ими ассимиляты лишь базипетально к многолетним частям и корневой системе. Таким образом жирующие побеги полезны для общего питания куста и запасания его корневой системы углеводами.
Интерес вызывают и следующие факты, установленные Коb1еt (1975, 1977). Первый из них касается того, что в конце лета (29/VIII) сила освещенности 800 1х (затененные листья) не обеспечивает отток ассимилятов из листьев. Несколько позже (13/IX) из затененных листьев при освещении 800 и 1800 1х обнаруживается отток. На этом основании автор считает, что для ассимиляции углерода листьев винограда нужна сила освещенности около 1000 1х. При периодическом освещении затененных листьев они начинают выносить ассимиляты.
Второй факт касается того, что листья винограда начинают выделять ассимиляты для потребностей других органов уже по достижении 30 % своей нормальной величины. Как только листовая поверхность достигает 50—70% окончательной величины, листья переправляют ассимиляты к другим органам. Лист отдает ассимиляты, как только те образуются, и передвигаются они со скоростью 30 cm/h. Перенос ассимилятов осуществляется главным образом сахарозой, однако в небольшом количестве принимают участие также глюкоза, фруктоза и другие неидентифицированные вещества.
В обобщающей работе „Migration des produits d’assimilation dans la vigne“ W. Koblet высказывает некоторые рекомендации, касающиеся практики виноградарства. Обломку побегов следует проводить на ранних этапах роста, сохраняя при этом нижние листья. Вскоре после цветения восьмой лист отправляет ассимиляты к основанию побега и молодым ягодам. В сентябре базальные листья II—III яруса не выносят ассимиляты, поэтому их можно удалять. Пасынки отдают большое количество углеводов ягодам винограда. Содержание сахаров в ягодах винограда определяется прежде всего фотосинтетической работой верхних листьев. Число пасынков оказывает положительное влияние на сахаристость ягоды. Для того, чтобы получить хороший урожай, нужно обеспечить 6—7 побегов и 12—18 соцветий на 1 m2 площади.