Весьма слабо исследовано влияние некоторых постоянных факторов при выращивании винограда — густоты посадки, подвоев, формировок, устройства опор. По мнению Тавадзе (1953, 1961) ассимиляция наиболее высока при двусторонней шпалерной формировке. В свою очередь Горбач (1958) устанавливает наиболее интенсивный фотосинтез при выращивании винограда на шпалерах с козырьком, при системе выращивания врасстил, а также с восточной стороны при всех системах формирования.
Наиболее высокую интенсивность фотосинтеза Тавадзе (1953, 1961) установил при сравнительно наименьшей густоте посадки — 3300 кустов на ha. Как увеличение числа кустов до 8800 и 13 000 на 1 ha, так и переход к формировкам типа Казенава приводят к понижению фотосинтеза. Это обстоятельство автор объясняет нарушением нормального соотношения между листовой поверхностью и корневой системой и ухудшением водного режима винограда,
Мишуренко и др.(1962) сообщают, что интенсивность фотосинтеза листьев сорта Серексия повышается при прививке на подвой 3309. При корнесобственной культуре или на подвоях Рипария Глуар интенсивность фотосинтеза ниже.
У сорта Шасла белая интенсивность фотосинтеза повышается при прививке на подвой 101—14 по сравнению с подвоем 1616. У сорта Рислинг фотосинтез выше при прививке на подвой 3309 по сравнению с 1616, а у Саперави — на подвое 1309 по сравнению с подвоем 1202.
По мнению Мамарова (1959) содержание хлорофилла в листьях в значительной степени повышается при прививке сортов Гымза и Мавруд. на подвои Рупестрис дю Ло и Кобера 5ББ по сравнению с прививкой на Шасла х Берландиери 41Б или же при корнесобственной культуре.
Сравнительно лучше изучено влияние некоторых агротехнических приемов — зеленых операций, орошения и удобрения на фотосинтетическую активность листьев.
Из исследований в этой области прежде всего следует отметить работу Кокиной (1937), изучавшей влияние прореживания листьев на интенсивность фотосинтеза. Наиболее высокую ассимиляцию Кокина установила при уменьшении числа листьев на 25 % и удалении боковых побегов — пасынковании. Удаление только боковых побегов (пасынков) или боковых побегов и 50—65% листьев незначительно повышает интенсивность ассимиляции. Характерно, что уменьшение листовой поверхности сильно увеличивает отток веществ.
По данным Кокиной трудно можно было бы определить, чему обязано повышение фотосинтеза — стимуляции вследствие поранения (травмы) или же лучшему освещению. Понижение активности фотосинтеза при сильном оперировании листовой поверхности автор объясняет сильным высушиванием листьев под воздействием света и ветра. О подобных результатах сообщает Берая (1977).
Никифорова (1959) также установила, что уменьшение числа листьев заметно повышает интенсивность фотосинтеза, вследствие чего количество СO2, усвоенного одним побегом, уменьшается незначительно. По данным автора сокращение листьев до 10—11 на побеге (примерно на 40% меньше, чем в контрольном варианте) повышает интенсивность фотосинтеза на 177— 214%.
Даже в варианте с оставлением лишь 5 листьев над гроздью в некоторых фазах вегетации установлено, что одним побегом усвоено наибольшее количество СO2. На основании этого автор приходит к выводу, что высокая энергия фотосинтеза, свойственная виноградной лозе, не используется полностью при обычных методах выращивания. Путем применения различных агротехнических приемов, связанных с рациональным регулированием величины листовой поверхности как куста в целом, так и отдельных побегов, пишет автор, можно повысить не только интенсивность фотосинтеза на единицу листовой поверхности, но и общую продуктивность растения.
Сравнительно более подробное исследование влияния прищипывания, пасынкования (удаления боковых побегов) и чеканки основных и боковых побегов на различных этапах вегетации на интенсивность фотосинтеза проведено Стоевым и Лиловым (1956). Результаты их исследования показывают, что применение зеленой обрезки приводит к временному повышению энергии фотосинтеза листьев виноградной лозы. Под влиянием прищипывания до цветения и во время цветения обнаруживается существенный сдвиг в сторону усиления фотосинтеза листьев, находящихся в секторе 4-го узла, до 145—244% против контроля. Значительное повышение энергии фотосинтеза (до 136—280%) отмечается и в вариантах, где прищипывание проводилось в разгар цветения. В то же время фотосинтез листьев в секторе 8-го узла, побега в тех же вариантах находится на уровне контрольных кустов, а в некоторых случаях даже значительно ниже. Все это показывает, что прищипка верхушек побегов виноградной лозы до цветения и в разгар цветения оказывает благоприятное воздействие на ход физиологических процессов одних лишь расположенных ниже листьев (в секторе грозди), не касаясь листьев в секторе 8-го узла.
Зеленые операции, применяемые позднее (ранняя чеканка), наоборот, усиливают фотосинтез расположенных выше листьев (в секторе 8-го узла до 180—225% против контрольных). В то же время в листьях зоны 4-го узла не отмечается усиления интенсивности фотосинтеза, за исключением случаев, где применялась сильная чеканка (на 12 листьев). Следует отметить также, что воздействие обрезки зеленых частей виноградной лозы на фотосинтетическую активность листьев, несмотря на некоторые отклонения, имеет хорошо выраженный временный характер.
Наибольший сдвиг к усилению фотосинтеза листьев отмечается к 6—10-му дню после применения обрезки. К 15-му дню интенсивность фотосинтеза под влиянием обрезки значительно ослабевает, хотя в некоторых случаях она все же выше, чем у листьев контрольных кустов.
Таким образом, воздействие обрезки зеленых частей на фотосинтез листьев виноградной лозы не статично, а динамично. Динамичность процесса выражается прежде всего в том, что воздействие носит временный характер, совпадая с приостановкой роста побегов, а также в том, что максимальный эффект наблюдается при постепенном поднятии побега кверху. Это, по-видимому, связано, с одной стороны, с зоной действия обрезки, а с другой — со степенью жизнедеятельности листьев, так как в течение вегетации жизнедеятельность старых листьев постепенно ослабевает.
Рябчун (1975) также указывал на усиление фотосинтеза под влиянием чеканки. Кондря (1962) установил, что после чеканки заметно повышалась интенсивность фотосинтеза.
Это повышение, по мнению автора, зависит от сорта (у Совиньон больше, чем у Муската белого), от интенсивности чеканки (при сильной чеканке больше, чем при слабой), от времени проведения операции. Несколько иные результаты получил Hernandez (1977).

Рис. 12. Интенсивность фотосинтеза листьев (mg CO2/dm2 на час) в секторе V и XVII узлов основного побега
1- под влиянием кольцевания; 2 — без кольцевания
Влияние на интенсивность фотосинтеза оказывает и кольцевание. Исследования Стоева с сотр. (1966) сорта Болгар показывают, что кольцевание подавляет фотосинтез листьев, причем это явление имеет временный характер — депрессия длится до восстановления связи проводящей системы в месте кольцевания.
Фотосинтез листьев на кустах, где не применялось кольцевания, всегда выше. В конце периода исследования, проведенного в 1964 г. (на 44—45-й день после кольцевания), разница небольшая, иногда даже фотосинтез выше там, где применялось кольцевание (рис. 12). На рис. 13 видно, что до кольцевания фотосинтез листьев подопытных кустов был почти одинаков. После кольцевания фотосинтез листьев окольцованных побегов значительно ниже, чем листьев неокольцованных побегов. Разница постепенно уменьшается и исчезает полностью лишь на 37-й день, на основании чего можно предположить, что к этому времени отток ассимилятов возобновляется.
Аналогичные данные получил также Hofacker (1978) — нетофотосинтез листьев окольцованных побегов гораздо ниже, чем листьев неокольцованных побегов. Уменьшение числа листьев до 75% сохраняет высокую интенсивность фотосинтеза остальных листьев почти до конца вегетации.
Исследования Мельника и Анисимовой (1958)показывают, что листья плодоносных побегов фотосинтезируют сильнее, чем листья бесплодных побегов. К концу вегетации, однако, фотосинтетическая активность листьев бесплодных побегов в значительной степени повышается.
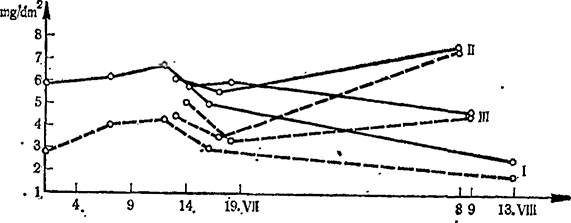
Рис. 13. Сезонная динамика интенсивности фотосинтеза листьев (mg СO2/dm2 на час) под влиянием кольцевания (пунктирная линия) и без кольцевания (сплошная линия) по среднесуточным данным за 1964 г.
I — листья 4—6-го узла; II — листья 10—12-го узла; III — листья 17-го узла
На основании этого авторы считают, что при выращивании винограда необходимо обеспечивать определенное соотношение между плодоносными и бесплодными побегами.
Тодоров (1961) также устанавливает, что листья плодоносных побегов фотосинтезируют интенсивнее по сравнению с листьями жирующих (порослевых) побегов. Содержание хлорофилла в них ниже, а содержание воды выше. Дыхание также интенсивнее у плодоносных побегов.
Как показали исследования Басанько, Горячевой и Рябчуна (цит. по Захаровой, 1964), значительное влияние на энергию фотосинтеза оказывает нагрузка побегами и гроздями. Различная нагрузка кустов побегами и гроздями изменяет как интенсивность работы единицы листовой площади, так и продуктивность работы куста в целом.
Наличие грозди на побеге способствует некоторому повышению интенсивности фотосинтеза. Осуществляется ускоренный отток продуктов фотосинтеза в пользу активного потребителя этих продуктов — грозди, что ведет к стимуляции усвоения СO2.
Чрезмерная нагрузка куста побегами и гроздями по мере увеличения листовой поверхности ведет к загущению и затенению части листьев и, следовательно, к ухудшению и относительному ослаблению ассимиляционной активности куста. Перегрузка куста чрезмерной листовой поверхностью, повышая затрату влаги и питательных веществ, поступающих из корней, может привести к ослаблению процессов фотосинтеза.
В силу указанных причин увеличение нагрузки до оптимума, по мнению авторов, содействует повышению фотосинтетической активности и продуктивности работы виноградного куста. Перегрузка, в свою очередь, снижает активность фотосинтеза и ухудшает состояние перегруженных кустов.
Влияние орошения на фотосинтез изучалось немногими авторами.
Исследования Стоева и Магрисо (1955) показали, что орошение в значительной степени повышает интенсивность фотосинтеза. Наиболее значительное усиление фотосинтетической активности листьев отмечается на 3— 8-й день после орошения. В некоторых случаях устанавливается повышение фотосинтеза в 2—10 раз по сравнению с фотосинтезом листьев на неорошаемых участках.

Рис. 14. Сопоставление интенсивности фотосинтеза (в g сухого вещества на m2) с величиной сосущей силы (at) и влажностью почвы (%)
1 — сосущая сила орошаемых кустов; 2 — сосущая сила неорошаемых кустов; 3 — фотосинтез листьев орошаемых кустов; 4 — фотосинтез листьев неорошаемых кустов; 5 — орошаемый участок; 6 — неорошаемый участок
По данным авторов существует положительная корреляция между интенсивностью фотосинтеза, с одной стороны, и сосущей силой и влажностью почвы — с другой. Чем ниже сосущая сила и чем выше влажность почвы, тем выше интенсивность фотосинтеза. Орошаемые кусты всегда показывают более низкую величину сосущей силы и более высокую интенсивность фотосинтеза. Иногда за незначительными изменениями влажности почвы следуют заметные изменения сосущей силы, что сказывается на интенсивности фотосинтеза (рис. 14).
Сосущая сила как показатель, характеризующий в некоторой степени энергетическое состояние воды, выражает физиологическую активность воды в клетках паренхимы орошаемых и неорошаемых кустов. Повышенное содержание воды в почве оказывает положительное влияние на фотосинтез в результате изменения осмотических свойств клеток ассимиляционной ткани до наступления заметных изменений в оводненности листьев.
По данным Кондо (1946) при нормальном водоснабжении в утренние часы интенсивность фотосинтеза примерно в два раза выше, а в дневные часы — около десяти раз выше по сравнению с фотосинтезом кустов, выращиваемых в условиях плохого водоснабжения. Достаточные водные запасы в почве, по мнению Кондо, устраняют депрессию в полуденные часы, вследствие чего листья отличаются довольно высокой ассимиляционной способностью в эти часы дня.
Весной и в начале лета, когда неорошаемые растения не испытывают еще недостатка воды в почве, интенсивность фотосинтеза у них немного отстает от интенсивности у орошаемых растений. С ухудшением водоснабжения общий жизненный тонус богарных растений падает, ухудшается аэрация листьев, в несколько раз снижается интенсивность фотосинтеза.
По данным Магрисо и Славчевой (1975а) между содержанием почвенной влаги в интервале с величины завядания до 70% от ППВ и интенсивностью фотосинтеза существует положительная корреляция. При влажности 70—100%, однако, корреляция не обнаруживается.
Подобные результаты получил и Hofacker (1976). По его данным понижение почвенной влаги вызывает снижение интенсивности фотосинтеза, причем при влажности почвы ниже 50—60 % различия достоверны. Выше этого интервала (100—60%) установлено незаметное уменьшение фотосинтеза.
Имеется ряд работ, в которых отмечается положительное влияние условий минерального питания на фотосинтез (Чанишвили, 1977; Корнейчук, П л а к и д а, 1975; Milosavlievic с сотр., 1977), а также влияние подвоя (Дзагнидзе, 1977; Тавадзе, Дзагнидзе, 1971, 1971а,
1972) и др. Без сомнения, такие исследования полезны. Несмотря на это, однако, приходится констатировать, что пока они не дают исчерпывающей информации о сложном влиянии минерального питания на фотосинтез, поэтому необходимо дальнейшее их проведение на более высоком методическом уровне.
Этим требованиям в известной степени отвечает работа Sautter (1971), поэтому мы вкратце остановимся на ней. Автор изучал влияние минерального питания на интенсивность фотосинтеза листьев одно-двухлетних растений сортов Арис, Рислинг и Сильванер, выращивавшихся в сосудах в теплице. Меняя соотношение и дозы минеральных веществ, автор получил следующие результаты.

Рис. 15. Влияние удобрения на интенсивность фотосинтеза (mg СO2/ dm2 на час) листьев сорта Рислинг
а — азотного удобрения на 1 растение; б — комбинированного удобрения NРК на интенсивность фотосинтеза листьев сорта Арис (g на 1 растение)
Во-первых, он установил, что между удобрением азотом и интенсивностью фотосинтеза существует непосредственная, тесная зависимость. В вариантах без удобрения или же с низкими дозами азота величина фотосинтеза невысокая — 2,68 ±0,186 mg CO2/dm2 на час. С повышением доз азотного удобрения повышается и интенсивность фотосинтеза до 4,22±0,10 mgCO2dm2 на час (рис. 15). Как видно из данных, после оптимума повышение дозы азота вызывает депрессию фотосинтеза; неодинакова также реакция разных сортов.
Подобное влияние на фотосинтез оказывает и калийное удобрение. Одновременное внесение возрастающих доз N, Р, К вызывает значительное повышение интенсивности фотосинтеза. Установлена также прямая связь между показателями фотосинтеза при различных условиях минерального питания и приростом вегетативной массы подопытных растений.
Почти аналогичные результаты получили Михайлова и Славчева (1976). Они установили, что увеличение доз N, Р, К в 2 раза повышает интенсивность фотосинтеза на 60% и листовую поверхность в 2,4 раза.