Влияние некоторых экологических условий на интенсивность фотосинтеза1
Исследования, посвященные проблеме влияния экологических факторов на фотосинтез листьев винограда, немногочисленны. При этом в большинстве случаев в них рассматривается влияние одного фактора внешней среды на фотосинтез и водный режим почвы (М агрисо, Славчева, 1975; Стоев, Магрисо, 1955; Gеis1еr, 1963; Hofacker, 1976), фотосинтез и солнечная радиация (Kriedemann и др., 1973) и т. д. Лишь в некоторых работах (Моторина, 1958; Kriedemann, 1968; Kriedеmann, Smаrt, 1971) имеются сведения о влиянии нескольких факторов на фотосинтез. Несмотря на это, в упомянутых исследованиях влияние факторов рассматривается независимо друг от друга.
В природных условиях, однако, факторы внешней среды оказывают влияние на фотосинтез в сложной взаимосвязи.
Если между факторами не было бы взаимодействия и они не оказывали бы влияния друг на друга, суммарный эффект был бы аддитивным на всех уровнях и процесс фотосинтеза можно было бы представить графически семьей параллельных кривых, исходя из формулы Y=f(A)+f(B) + ...+ const. (f—функция изучаемого фактора). В действительности же факторы внешней среды оказывают влияние друг на друга, кривые непараллельны и процесс фотосинтеза можно представить формулой Y=f(A).f(B). . . к или же другими соотношениями, отвечающими более точно данному типу взаимодействия (X и т, 1972).
Сведения об оптимальной температуре фотосинтеза листьев винограда весьма разноречивы. По данным Bosian (1964, 1968) оптимальной является температура 30 °С, причем, по его мнению, ведущий фактор интенсивности фотосинтеза — свет. Kriedemann (1968, 1975), Kriedemann, Smart (1971) и Кriеdеmann, Lеnz (1972) считают, что температурный оптимум листьев винограда — 25°С в тепличных условиях и 30°С— при выращивании в открытом грунте. Интенсивность фотосинтеза быстро снижается при 35 °С, доходя до 0 между 45 и 50°С. Если гидратура листьев поддерживается на высоком уровне, ассимиляция СO2 обнаруживается при температуре 48—49° на протяжении 3 h. Выше 50°С установлен обратный ток СO2, и наступает обесцвечивание отдельных листьев — листья гибнут.
1 Материал разработан К. Д. Стоевым и Т. Славчевой.
По данным Солэджану (1962) листья виноградной лозы, оставленные в течение краткого времени (5 min) в условиях высокой температуры — до 50°С, сохраняют свою способность поглощать СO2. Homey (1973) считает, что максимальная интенсивность фотосинтеза наблюдается в температурном интервале 20—28 °С. По его мнению возможно даже некоторое сужение интервала в границах 25—28 °С, а при температуре выше 30°С интенсивность фотосинтеза быстро падает. Несколько более низкий температурный оптимум установил Sautter (1971) для растений, выращиваемых в теплицах. Для них повышение температуры за 20 °С (20—30o С) приводило к понижению интенсивности фотосинтеза до 40 %. Резкое снижение интенсивности фотосинтеза при температуре выше оптимальной обнаружил и Р1аntеfо1 (1950).
Таким образом, виноград проявляет значительную толерантность к температурному фактору. По неопубликованным данным Т. Славчевой при температуре воздуха 8—10°С интенсивность фотосинтеза значительна (5—6 mg CO2/dma на час). Она установила также, что изменения температуры в кюветах в пределах 17—33 °С вызывают незначительные изменения интенсивности фотосинтеза, а отклонения температуры выше 33 и ниже 17 °С вызывают более значительные изменения в интенсивности фотосинтеза.
Влияние температурного фактора на интенсивность фотосинтеза менее заметно, если свет является лимитирующим фактором, что находится в соответствии с установленным для других растений (Либберт, 1976).
Обнаруженные авторами различия, по всей вероятности, будут несколько сглажены, если принять во внимание, что иногда данные о температуре относятся к атмосфере, иногда к температуре листа.
Исследуя влияние температуры на фотосинтез, Моторина (1958) устанавливает, что температурный оптимум зависит в значительной степени от сорта. У сорта Альфа фотосинтез повышается до 20—25 °С, после чего заметно падает (исследование проведено при освещенности более 20 000 1х и относительной влажности воздуха 65—75%). Понижение фотосинтеза у сорта Альфа с точки оптимума в направлении низкой температуры — с 24 до 14°С происходит плавно и составляет около 3 mg CO2/dm2 на час. Более резко понижается фотосинтез (около 4 mg) при повышении температуры выше оптимума — с 24 до 30°С. Аналогичная реакция к температуре установлена и у сорта Амурский. Сорт Черный сладкий, однако, реагирует положительно на более высокие температуры — 28—30°С. При высокой влажности воздуха температура в 32 °С не оказывает угнетающего влияния.
Установленный у сортов Альфа и Амурский оптимум не соответствует оптимальной температуре для других растений, выращиваемых в условиях средней географической широты. Эти два сорта отличаются высокой морозоустойчивостью и более коротким вегетационным периодом. Третий сорт (Черный сладкий) отличается более длинным вегетационным периодом и значительно меньшей морозоустойчивостью. Как более теплолюбивый, он обладает способностью переносить повышенные температуры. Отмеченные различия, по мнению автора, соответствуют экологическим условиям, в которых произошло формирование их биологических качеств.
Из внешних факторов свет — основное условие для осуществления фотосинтеза, поэтому ряд исследователей уделяет внимание изучению влияния этого фактора.
По данным Gеis1еr (1963а) компенсационные точки света варьируют в пределах 300 и 15001х, причем они неодинаковы для разных сортов и видов — наиболее низкая граница специфична для V. riparia. а наиболее высокая — для V. labrusca. Величина насыщенности света также неодинакова и по данным Gеislеr (1963а) находится в пределах 10 000 lx (V. riparia) и 30 000 1х (V. rupestris). Эти особенности видов автор объясняет характером солнечной радиации их местообитаний.
По данным других авторов (Кriеdеmаnn, 1968; Kriedemann, Smart, 1971) световая насыщенность фотосинтеза винограда в теплицах или же в открытом грунте, но в тени, наступает при 35 000 1х. Для виноградных кустов, выращиваемых в полевых условиях, пункт световой насыщенности определяется в пределах 40 000—60 000 1х в зависимости от того, где проводились измерения — в полевых условиях или же в лаборатории. При этом величина фотосинтеза винограда в теплицах или в открытом грунте почти одинаковая — до 10х104 erg/s. cm2, т. е. 27 000 1х, после чего кривые расходятся. Световая интенсивность выше 15х104 erg/cm2.s (40 000 lx) вызывает небольшое повышение фотосинтеза листьев растений в открытом грунте, однако подавляет или не вызывает дальнейших изменений фотосинтеза листьев в тепличных условиях (Kriedemann, 1968а; Kriedemann, Smart, 1971). Это показывает, что виноградная лоза подобно другим растениям в природных условиях использует свет не вполне продуктивно.
Установлено также, что условия выращивания создают временную адаптацию при перемещении виноградных растений из теплицы в открытый грунт. Так, через пять дней после выноса растений из теплицы интенсивность фотосинтеза снижается на 50% (Kriedemann, 1968а), в дальнейшем такого эффекта не устанавливается. Обратное перемещение растений не вызывает реципрокных изменений в максимальных величинах интенсивности фотосинтеза.
Kriedemann и Smart (1971) установили, что когда освещенность ограничена и отсутствует диффузный свет, фотосинтез определяется косинусом угла падающего света. При полной освещенности косинусная зависимость нарушается, и если свет проходит даже параллельно расположению листьев, интенсивность фотосинтеза снижается лишь на 50%.
Листья винограда обнаруживают также в некоторой степени и гелиотропизм — это явление установлено у сильнорослых растений в условиях орошения. В новейших исследованиях Smart (1974) устанавливает, что в компактных кронах виноградных кустов фотосинтез осуществляется главным образом наружными листьями, которые абсорбируют прямо падающий свет, в то время как величина диффузного света внутри кроны очень низка и подавляет усвоение углекислоты. Однако диффузным светом в менее плотных кронах не следует пренебрегать. Квантовый эффект фотосинтеза улучшается при прерывании светового потока каждые 1,2 s на 0,6 s. По данным Kriedemann и др. (1973) листья винограда, находящиеся в глубине кроны, в состоянии компенсировать затраты на дыхание, если диффузная радиация находится близ компенсационной точки (675—1350 1х), при условии, что 1% их поверхности периодически получает освещение насыщающей плотности.
Сведения о световом оптимуме виноградной лозы встречаются и у других авторов. По данным В о s i а η (1964) в кондицированных кюветах оптимальная освещенность для фотосинтеза винограда составляет 60 0001х. В условиях пустыни Негев, Schulze и др. (1972) устанавливают, что световая насыщенность для V. vinifera в утренние часы наступает при 60 000 1х. Повышение интенсивности света до 90000 1х вызывает незначительный рост интенсивности фотосинтеза (0,5 mg CO2/dm2 на час) при максимальных величинах (6,5 mg CO2/dm2 на час). Эти данные авторы рассматривают как приспособительную реакцию винограда к условиям выращивания (интенсивность света 60—100 klx, температура воздуха 27—38 °С и влажность атмосферы ниже 40%).
В отличие от этих данных Homey (1973) считает 20 000 1х оптимальной освещенностью, установленной при температуре 25 °С, 60% влажности воздуха и 75—80% от ППВ. Совершенно иной характер имеют сведения Гриненко и Фоменко (1977). По их данным световая насыщенность не наступает и при 120—140 klx при условии, что температура и влажность воздуха находятся в оптимуме (27,9—30,5 °С и 66—78%). Снижение влажности воздуха до 42% при сохранении указанной температуры смещает порог насыщенности света до 50 000—60 000 1х, а при повышении температуры до 32—34 С оптимальная сила света оказывается в пределах 30 000—50 0001х.
Подводя итоги данным исследований по установлению оптимальной интенсивности света, нельзя не видеть значительной разноречивости, которая не может быть объяснена лишь методикой работы. Как нам представляется, большое значение имеют сортовые и видовые особенности винограда, условия выращивания и сочетание экологических условий при экспериментировании. Поэтому проблему оптимальной интенсивности света для фотосинтеза листьев винограда не приходится считать решенной, и ее выяснение требует дальнейших исследований.
Исследования Моториной (1958) по выяснению влияния интенсивности освещения показали, что с усилением света до 20 000 1х, а для отдельных сортов и до 30 000 1х, интенсивность фотосинтеза повышается. Усиление света выше этого предела, по мнению автора, не отражаемся на фотосинтетической активности листьев. К аналогичным выводам приходят также Басанько и Горячева (1964).
Внимательное рассмотрение данных М. В. Моториной, однако, показывает, что у сорта Черный сладкий при изменении освещенности с 20 000 до 100 000 1х и влажности воздуха 50—60% обнаруживается повышение интенсивности фотосинтеза (2 mg CO2/dm2 на час), которым автор пренебрегает.
Некоторые авторы считают, что чем более к северу выращивается растение, тем больше оно нуждается в свете — необходимость в свете возрастает по мере понижения температуры. Расположение листа винограда в тени ведет к понижению интенсивности фотосинтеза. Как установил Mfiller-Thurgau (цит. по Мержаниану, 1939), лишь хорошо размещенные по отношению к освещению листья способны к максимальной ассимиляции. Исследования Kriedemann (1968), однако, показали, что при значительном снижении интенсивности света фотосинтез сохраняет довольно высокую интенсивность. Эксперимент автора заключался в следующем: полностью развитые листья сорта Султанина были поставлены в кювету, в которую свет проходил через отверстие диаметром всего в 3 cm и освещал лишь 8,5 % поверхности листа. Несмотря на это, продолжительное воздействие пучка света интенсивностью 15 х 104 erg/s. cm2 обеспечивало 25 %-ную активность фотосинтеза по сравнению с листом, полностью освещенным. Прохождение пучка света с перерывами до 6 раз и продолжительностью света 0,02 s обеспечивало поглощение СO2 в размере выделенной дыханием углекислоты.
При постоянном освещении интенсивность фотосинтеза зависит от содержания влаги в воздухе — с увеличением относительной влажности фотосинтез повышается. Исследования Моториной (1958) показывают, что понижение относительной влажности воздуха с 70 на 50% угнетает весьма резко интенсивность фотосинтеза.
Воsiаn (1964) и Alleweldt (1963) установили, что ассимиляция СO2 понижалась по мере прогрессивного снижения насыщения листьев водой. Действие засухи, однако, смягчается подвоем. Понижение фотосинтеза у новой формы (Aris), устойчивой к паразитическим грибам и филлоксере и выращиваемой на сухой почве (50% ППВ), составляет лишь 0,92 mg CO2/dm на час по сравнению с фотосинтезом листьев сорта, выращиваемого на влажных участках (80% ППВ). У гомопластов и прививки понижение достигает 1,20 mg CO2/dm2 на час. Эффективность подвоя С 3309 выражается в 1,08 mg CO2/l00 dm2 на час. Наконец, использование подвоя Кобера 5ББ приводит к различию в 0,28 mg CO2/dm2 на час при выращивании на влажных и сухих участках (Alleweldt, 1963).
К riedemann и Smart (1971) и Kriedemann (1975) установили, что интенсивность фотосинтеза постепенно снижается, если водный потенциал листа выше —5 bar и полностью приостанавливается, когда его величина превышает —15 bar. Восстановление водного статуса листа наступает непосредственно после орошения, в то время как восстановление нормальной фотосинтетической активности наступает значительно медленнее — после 5 дней. Неодновременное восстановление влажности листьев и фотосинтеза после стресса вызвано действием абциссиновой кислоты. Под влиянием обезвоживания тканей листа наступает чрезмерное накопление абциссиновой кислоты (иногда в 44 раза больше нормального содержания), которая вызывает закрытие устьичных щелей и задерживает восстановление их нормального состояния на некоторое время после орошения.
Фотопериод, определяющий рост побегов и ряд других физиологических процессов (Alleweldt, 1963), не оказывает непосредственного влияния на интенсивность фотосинтеза. В условиях равномерного во все дни освещения и при циклах различной продолжительности можно наблюдать прирост сухого вещества виноградных кустов в период вегетативного роста, как и в дни с длительным фотопериодом: это нарастание основывается на более полном использовании света (Alleweldt, 1963).
Углекислота — основной субстрат фотосинтеза, и от ее содержания зависит в значительной степени интенсивность фотосинтеза. В отличие от других растений (Koch, 1969; Friedrich, 1976) влияние концентрации СO2 на фотосинтез винограда очень мало исследовано. По данным Kriedemann (1968) для вполне развитых листьев и в тепличных условиях с повышением концентрации СO2 до 300 ppm (более высокие концентрации СO2 не изучались) при освещенности 65 000 1х и t=28,5СС фотосинтез нарастает линеарно.
Влияние повышенных концентраций СO2 исследовано при проведении термотерапии как средства повышения устойчивости растения к высоким температурам (Кriеdеmаnn и др., 1976). Полученные данные указывают на благоприятное воздействие повышенных концентраций СO2 на рост побегов и, особенно, корневой системы, а также на усиление эффективности водопотребления (в 4 раза). Это находит свое объяснение в том, что при высоких температурах транспирация сокращается на 50% из-за частичного закрытия устьичных щелей, а фотосинтез возрастает в два раза. Как правило, листья кустов, развивавшихся в камерах, обогащенных СO2, реагировали сильнее на повышенные концентрации углекислоты. Наблюдаемый феномен авторы считают признаком фотосинтетической адаптации к обогащению СO2 для растений, развивавшихся в условиях высоких температур (37—40°С). При этом повышенная фотосинтетическая деятельность сопровождается уменьшением фотодыхания, что также благоприятствует усилению роста при повышенных температурах.
Заслуживают внимания еще две работы, рассматривающие влияние концентрации СO2 на интенсивность фотосинтеза. Plantefol (1950 ) установил, что повышение концентрации СO2 до 0,1% (в 3 раза) вызывает крутой подъем в интенсивности фотосинтеза. Дальнейшее повышение СO2 до 0,3% усиливает фотосинтез, однако это происходит очень медленно и непропорционально. Тем временем Binet и Brunei (1967—1968) показали, что эффективность концентрации во многом зависит от освещенности: при 6000 1х максимальная интенсивность фотосинтеза обнаруживается при 1 % СO2, при 20 000 1х оптимальная концентрация СO2 уже 1,5%, а при 120 000 1х максимальная интенсивность фотосинтеза выявляется при 2% СO2.
Влияние других экологических факторов (географической широты, высоты над уровнем моря, состава почвы) на интенсивность фотосинтеза недостаточно выяснено. В литературе встречаются отдельные сведения о том, что местообитание виноградной лозы оказывает некоторое влияние на интенсивность фотосинтеза (Чрелашвили, 1959). Не установлено с достаточной убедительностью влияния повышенных количеств кальция в почве и появления хлороза на фотосинтетическую активность листьев (Мазанашвили, 1954).
Исследования, проводившиеся в Институте виноградарства и виноделия в г. Плевен на протяжении трех лет (1975—1977), имели целью изучение интенсивности фотосинтеза по возможности на фоне различных сочетаний нескольких факторов (Славчева, Стоев, 1978; Стоев, Славчева, 1979). В качестве опытного материала использовались однолетние и двухлетние растения винограда сортов Ркацители и Каберне Совиньон, которые выращивались в тепличных условиях в сосудах типа Вагнера.
Измерения проводились в открытом грунте или в частично кондиционированных камерах. Интенсивность фотосинтеза определяли инфракрасным газоанализатором — URAS-2.
В наших исследованиях изучалось совместное влияние освещенности, температуры, водного режима и концентрации углекислоты на интенсивность фотосинтеза листьев винограда. Здесь сообщаются сведения о световых кривых при разных сочетаниях водного режима, температурного фактора и концентрации углекислоты. В качестве источника света использовали люминесцентные лампы типа ЕС 40 W и 120 W и ртутные лампы 400 W.
На рис. 8 представлена зависимость фотосинтеза от освещения при разной влажности почвы.

Рис. 8. Световые кривые (интенсивность фотосинтеза, mg CO2/dm2 на час) при различной влажности почвы
1 — выше 75% от ППВ; 2 — 65—75% от ППВ; 3 — 45—55% от ППВ; 4 — 35—45% от ППВ
Как видно из данных, при влажности почвы 35—45% от ППВ выявляется отрицательный баланс углекислоты, который повышается незначительно с увеличением освещенности. При повышении влажности почвы до 45—55% от ППВ интенсивность фотосинтеза повышается примерно до 13,5.103 lx, после чего обнаруживается тенденция к снижению. При влажности почвы до 65—75 % и выше 75 % от ППВ длина линейных участков увеличивается до 17.103 1х.
Исследования интенсивности фотосинтеза, проведенные в 1977 г., когда сосуды помещались вне теплицы, дали почти такие же результаты, что и в лабораторных условиях. При очень низкой влажности почвы (35% от ППВ) осуществляется процесс фотосинтеза, однако при 13 000—30 000 1х его интенсивность резко снижается (около 0,4 mg CO2dm2 на час) и полностью приостанавливается при 40.10 1х. При влажности почвы 50 и 80% от ППВ интенсивность фотосинтеза обнаруживает нормальный ход до 20.10 — 30.10-1х, после чего наступает спад.
Установлено также взаимное влияние температуры воздуха, освещенности и влажности почвы на фотосинтез. На рис. 9 а, б представлен суточный ход фотосинтеза, температуры и освещенности при влагообеспеченности 35 и 70% от ППВ. Экспериментальные данные показывают, что в обоих случаях кривая интенсивности фотосинтеза является как бы зеркальным отражением кривой суточных изменений температуры, с той лишь разницей, что температурный фактор имеет неравнозначное значение при разных условиях влагообеспеченности. При влажности почвы 35 % от ППВ минимальный фотосинтез установлен лишь при температуре ниже 20 С и более низких значениях освещения.

Рис. 9. Ход фотосинтеза (1) при 70% от ППВ (а), и 35% от ППВ (б), в связи с изменением температуры (2) и освещенности (3)

Рис. 10. Световые кривые
а — при неизменной концентрации 0,03—0,04 об. % СO2 и температуре воздуха: 1—18—21°С; 2 -27 -33°С; 3— 36—39°С и 4 — 42—45СС, б— при высоких концентрациях СО и температуре воздуха: 5 — 25—29 С; б — 21—25°С: 7— 17 21 С и 8 — 13—17°С
По достижении температурным фактором 20—23 С и освещенности 48.103 1х обнаружен отрицательный баланс углекислотного обмена, что свидетельствует о превалировании дыхания над фотосинтезом. При влажности 70% от ППВ усвоение углекислоты установлено при любых значениях воздуха, причем минимум (2 mg CO2/dm2 на час) наблюдается лишь при 36°С и освещенности более 80.103 1х. Таким образом, оптимальный водный режим сказывается на ходе фотосинтеза при высоких значениях температуры и освещения.
На рис. 10 представлены световые кривые фотосинтеза при неизменной концентрации СО, (0,3—0,4 об. %), но при разных температурных условиях. Из этих данных следует, что наиболее высокие величины интенсивности фотосинтеза обнаруживаются при температуре воздуха в пределах 21—27°С, несколько ниже они при температуре 27—33 °С и еще более снижаются при температуре 36—39 и 42—45°С. Вывод, вытекающий из этих данных, сводится к тому, что чем выше температура оптимального предела, тем больше она становится лимитирующим фактором, что установлено и многими другими авторами ранее (Моторина, 1958; Kriedemann, 1968; Кriеdеmann, Smart, 1971). Характер влияния светового фактора при разных температурных условиях сохраняется.
При высоких концентрациях СO2 максимальные величины интенсивности фотосинтеза установлены в температурном интервале 21—25 °С. Статистическая обработка данных при температурных интервалах через 2°С показала, что оптимальной для фотосинтеза в условиях высоких концентраций СO2 является температура воздуха в пределах 25—27 С. Вместе с тем повышение концентрации СO2 заметно повышает потребности в большей интенсивности света — до 41.103 1х кривые приобретают сугубо линейный характер. При более низких концентрациях СO2 (0,03 %) оптимальная освещенность находится в пределах 20.103—30.103 1х.
Из данных табл. 2 видно, что при освещенности выше 10.103 1х ведущим фактором, воздействующим на величину фотосинтеза, является концентрация СO2. При освещенности 16,5.103—17,5.102, 27.103—30.103 и 36,5.102—38,5.102 lx увеличение концентрации СO2 с 0,04 (A0,04) до 0,07, 0,08, 0,09 и 0,10 об.% (Ас) приводит к повышению интенсивности фотосинтеза в 1,5—2 раза.
При концентрации СO2 0,1%, но при более высоких значениях света в условиях оптимального увлажнения получены очень высокие величины интенсивности фотосинтеза (табл. 3). Это показывает, что факторы взаимосвязаны и что любое изменение одного фактора приводит к изменению влияния другого фактора.
Загрязнение природной среды вызвало необходимость изучения его влияния на интенсивность фотосинтеза листьев винограда. В связи с этим Добрева с сотр. (1974) изучали влияние осадки пыли в цементной промышленности на работу листьев виноградной лозы. Для этой цели они выбирали подходящие подопытные листья III—IV, X и XV яруса побега (снизу вверх), которые обладали одинаковой интенсивностью фотосинтеза. На следующий день на 50% подобранных листьев наносили цементную пыль в среднем по 200—700 mg /dm2 и определяли динамику поглощения СO2 инфракрасным газоанализатором. Представленные данные на рис. 11 убедительно показывают, что цементная пыль снижает фотосинтетическую активность листьев винограда 3—4-го узла на 15—45%1 . Особенно чувствительны молодые листья винограда (10—15-го узла).
1 Для краткости приводится лишь часть полученных данных.

Таблица 2
Изменение интенсивности фотосинтеза при повышенных концентрациях СO2 (0,1°/о), высоких значениях освещенности
Таблица 3
| Освещенность, 103 1x | Интенсивность фотосинтеза, mg CO2/dm2 на час | Температура воздуха, °С | Влажность воздуха, |
10 | 25,72 | 28,3 | 78 |
14 | 33,36 | 28,3 | 78 |
20 | 49,81 | 28,3 | 78 |
32 | 53,94 | 21,2 | 91 |
42,5 | 65,12 | 21,2 | 91 |
46 | 79,38 | 25,2 | 91 |
41 | 72,85 | 25,5 | 88 |
41 | 62,51 | 21,2 | 91 |
По данным Jonescu, Elvira (1972) атмосфера, насыщенная SO2, пылью Pb и другими веществами, вызывает морфологические изменения в устьичном аппарате листьев растений и уменьшение числа нормально функционирующих устьичных щелей, что, в конечном итоге, создает предпосылки для снижения интенсивности фотосинтеза. Huber (1971) отмечает, что цементная пыль, входя в соприкосновение с влагой атмосферы, образует твердую оболочку на поверхности листьев растений.
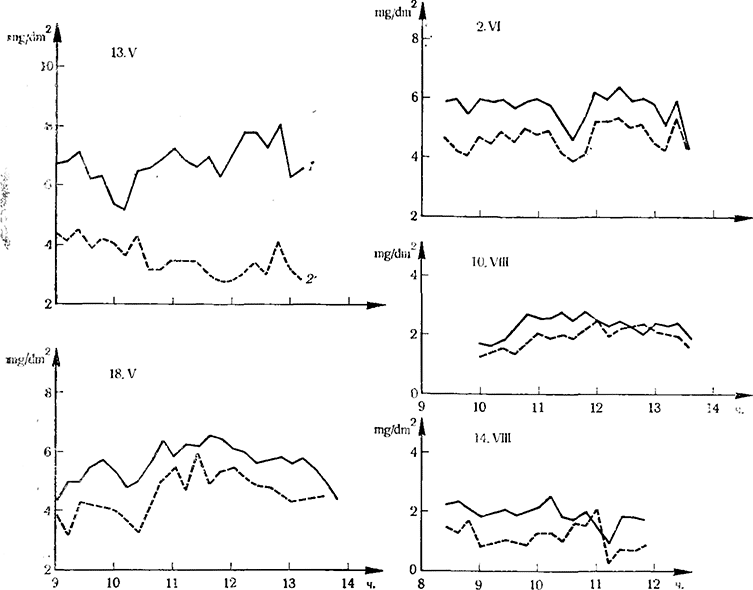
Рис. 11. Интенсивность фотосинтеза листьев винограда (mg CO2/dm2 на час), обработанных цементной пылью
1 — контроль; 2 — обработанные цементной пылью
Известняк вызывает также некроз листьев.
Как сообщают Thompson и Cats (1970) (цит. по Линник, 1971) вредное влияние оксидантов (окислов азота и некоторых углеводородов) можно устранить, применяя т. наз. антиоксиданты — дифенилендиамин (ДФФД) и Дуст. Влияние антиоксидантов выражалось в увеличении веса листьев, содержания хлорофилла, среднего веса и сахаристости ягод.