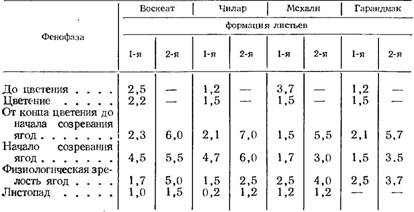
Из исследований, посвященных дыханию листьев, представляет интерес работа Н. М. Сисакяна, И. А. Егорова, Б. Л. Африкяна (1948) (табл. 82).
Из табл. 82 можно сделать следующие выводы.
Активность дыхания листьев всех изучаемых сортов винограда к листопаду снижается. У листьев второй формации (более молодые) она в несколько раз (в три и более) выше, чем у листьев первой формации (более взрослые). Максимум активности дыхания листьев второй формации приходится на период после цветения, в то время как у первой — на начало созревания (сорта Чилар и Воскеат). У сортов Мсхали и Гарандмак максимум дыхания листьев первой формации сдвигается на более поздний период, что можно объяснить более длинным вегетационным периодом у этих сортов.
Таблица 82
Дыхание листьев различных сортов винограда (CO2 в мг/ч на 50 см3)
Авторы отмечают, что не всегда имеется прямая зависимость между изменениями в интенсивности дыхания и активности ферментных систем. Обнаруженные различия в активности окислительно-восстановительных ферментов и биоактивных соединений, по мнению авторов, можно объяснить наличием у винограда различных дыхательных систем, та или иная из которых превалирует в различные фазы развития. Авторы отмечают, что в листьях винограда, как и в листьях яблони (Сисакян, Рубин, 1944), полифенолоксидазная и пероксидазная системы активнее начинают действовать лишь в поздние фазы развития. Активность дыхания и работа ферментов больше зависят от условий среды и возраста того или иного органа, чем от сорта. Условия агротехники могут вызывать значительные изменения в физиолого-биохимических процессах живой клетки. Определенный интерес в связи с этим имеют исследования Б. И. Библиной (1956), в которых изучалось влияние нагрузки куста побегами на интенсивность дыхания листьев при разных площадях питания растений. (Работа выполнена в условиях Молдавии, сорта Сенсо и Шасла.)

Рис. 67. Интенсивность дыхания листьев сорта Шасла:
1, 2— площадь питания 1,25x2,25 м, 20 (л и 40 (2) побегов на куст; 3, 4 — площадь питания 2,25х2,25 м, 24 (3) и 61 (2) побег, на куст
Результаты исследований показали изменение интенсивности дыхания листьев по фазам вегетации. Наиболее высокая интенсивность дыхания наблюдается в период массового цветения, к началу созревания ягод она снижается и затем в процессе созревания их несколько повышается.
В течение всего вегетационного периода интенсивность дыхания листьев у сорта Шасла выше, чем у сорта Сенсо. У первого значительно слабее выражены различия по этому показателю между отдельными вариантами площади питания и нагрузки побегами на куст (рис. 67).
Перегрузка побегами ослабляет дыхание, особенно при меньшей площади питания, а активность каталазы при этом в большинстве случаев усиливается в обоих вариантах площади питания. Ход изменений активности каталазы листьев часто не совпадает с ходом изменений дыхания. У сорта Шасла активность каталазы выше, чем у сорта Сенсо.
В другой работе Б. И. Библиной (1963) изучалось изменение содержания аскорбиновой кислоты как фактора, регулирующего превращения сахарозы в окислительно-восстановительных процессах и в азотном метаболизме растительной клетки, и дыхание (Рубин и Арциховская, 1937; Михлин, 1936).
Исследование интенсивности дыхания винограда в условиях Московской области у разных сортов в 1952—1953 гг. проведено М. В. Моториной (1958).
Из трех сортов винограда — Альфа, Черный сладкий и Амурский — первый отличался наибольшими величинами дыхания. В среднем интенсивность дыхания изменялась от 0,86 до 5,19 мг CO2 на 1 дм2/ч. В июне и июле расход органического вещества на дыхание составил 50—60% образовавшегося при фотосинтезе, в конце вегетации — 20,6—14,6%. У сорта Амурский максимум дыхания достигал 11,1 мг CO2 на I дм2/ч; в конце вегетации дыхание снизилось до 1 мг. В июле расход на дыхание у этого сорта составил 74—80% фотосинтеза. В конце июля и в августе фотосинтез резко снизился и отношение дыхание : фотосинтез осталось высоким — 30%. Средняя величина дыхания листьев у сорта Черный сладкий самая низкая — максимум 5,3 мг, 30—35% фотосинтеза в первой половине вегетации, 21% в июле и 10,9% в сентябре.
Как видно из приведенных данных, дыхание листьев винограда в условиях Московской области представлено довольно значительными величинами. Расход на дыхание по отношению к фотосинтезу составил в среднем 30—40%. При этом траты на дыхание большие в начале цветения, как и в более южных местах произрастания винограда. К концу вегетации они значительно уменьшаются и доля продуктивности фотосинтеза увеличивается. Уменьшение дыхания листьев во второй половине вегетации объясняется их возрастом.
По данным автора, повышение интенсивности дыхания и траты на дыхание при высокой интенсивности фотосинтеза находится в прямой связи с энергией роста растения. Сорт Альфа характеризуется большей силой роста, более продолжительным периодом активного фотосинтеза, повышенной интенсивностью дыхания и большими тратами на дыхание по отношению к фотосинтезу, чем сорт Черный сладкий. Сорта Альфа и Черный сладкий меньше отличаются по интенсивности дыхания, чем по отношению дыхания к продуктивности фотосинтеза. По мнению автора, для правильной оценки значения величины дыхания наряду с ней следует учитывать также и отношение дыхание : фотосинтез. Высокое отношение дыхания к фотосинтезу у сорта Альфа не сопровождается снижением общей продуктивности фотосинтеза, так как с возрастанием дыхания увеличивается энергия роста, что приводит к большему развитию ассимиляционной поверхности. Виноградное растение в условиях Московской области развивает высокую интенсивность дыхания, которая неодинакова у разных сортов винограда и превышает величины дыхания в других местах его произрастания.
Так, в условиях Эстонской ССР, по данным Мийдла (1961), на дыхание винограда расходуется в среднем 4—15% образовавшегося при фотосинтезе органического вещества.
Измерение дыхания в течение суток показало, что от 0 до 12 ч величина дыхания практически не менялась и составляла с 23 по 29 июня 1957 г. 0,60—0,61 мг CO2 на 1 г сырого вещества в 1 ч и 0,40—0,41 мг CO2 на 1 дм2 листа в 1 ч. Максимум дыхания приходился на дневные часы (12—16) и составлял 0,78 мг CO2 на 1 г сырого вещества в 1 ч и 0,53 мг CO2 на 1 дм2 листа в 1 ч. Минимум дыхания в течение суток наблюдался с 16 до 24 ч, 0,40—0,52 мг CO2 на 1 г сырого вещества в 1 ч и 0,27—0,35 мг CO2 на 1 дм2 листа в 1 ч.
Зависимость дыхания от возраста листьев та же, что и в других местах произрастания винограда. Так, в условиях холодных парников в июне дыхание молодых листьев составляло 0,63, среднего возраста — 0,36 и старых — 0,32 мг СС*2 в 1 ч.
Снижение интенсивности дыхания листьев во второй половине вегетации, как общее явление для разных мест произрастания и объясняемое не только возрастом, но и изменением внешней среды, у сортов и видов винограда, более приспособленных к условиям Эстонской ССР, выражено отчетливее, чем у менее приспособленных. В связи с этим, по мнению Мийдла, по интенсивности дыхания можно судить о степени приспособленности сортов винограда к конкретным условиям выращивания.

Рис. 68. Динамика интенсивности дыхания листьев сортов винограда различных сроков созревания (по Марутян, 1957):
1, 2 — ранние сорта (соответственно Сатени белый и Араксени белый): 3, 4 — поздние сорта (соответственно Арарати и Астамашк)

Рис. 69. Интенсивность дыхания и изменение содержания аскорбиновой кислоты в листьях винограда:
1— интенсивность дыхания; 2 — содержание аскорбиновой кислоты
В этом отношении интересно поведение раннеспелых и позднеспелых сортов винограда. Отдельные этапы жизнедеятельности у ранних сортов начинаются несколько раньше, чем у более поздних. В зависимости от условий местопроизрастания эти различия в начале вегетации могут почти отсутствовать. Однако позже все процессы у ранних сортов проходят быстрее и при большей активности, чем у позднеспелых (рис. 68).
Мооритс (1963) установил, что сезонные изменения содержания аскорбиновой кислоты в листьях винограда и интенсивность их дыхания изменяются параллельно (рис. 69). Первый максимум по обоим показателям наблюдается в фазе цветения; в период роста ягод наступает некоторое снижение, а после созревания ягод — новое повышение. Под влиянием повышенной температуры (33°С) интенсивность дыхания сильно возрастает, при этом содержание аскорбиновой кислоты снижается.
Работ, затрагивающих изменения интенсивности дыхания под влиянием удобрений, немного.
Работа, проведенная К. М. Плотниковой (1959) в условиях Киевской области в 1956 г. на сорте винограда Шасла, включает наблюдения за изменениями активности дыхания листьев под влиянием микроэлементов (Мп, В) на фоне минеральных удобрений (NPK). Удобрения вносили в почву в жидком виде перед цветением растения.
Интенсивность дыхания листьев с 13 июня по 9 июля (цветение и рост ягод) в варианте NPK снизилась. Подъем наблюдался в конце июля при подготовке ягод к началу созревания, а затем снова наступило снижение. В варианте с микроэлементами интенсивность дыхания на протяжении периода наблюдений была выше, чем в варианте NPK. Наибольшее усиление дыхания отмечено в варианте КРК+Мп + В и несколько меньшее в варианте NPK+Mn. Такое же влияние оказали микроэлементы на повышение содержания в листьях хлорофилла, однако эти изменения по фазам вегетации не совпадают с изменениями интенсивности дыхания.
Т. Мартин и его соавторы (1966) изучали влияние микроэлементов на интенсивность дыхания листьев сорта Рислинг итальянский при орошении в условиях Румынии. Увеличение дыхания через 3 дня после первого полива (период цветения) в среднем составило 13,5%, через 6 дней — 11,7% и через 3 дня после второго полива (период созревания) —8,4% по сравнению с контролем. Наибольшее увеличение интенсивности дыхания через 3 и 6 дней после первого полива наблюдалось в варианте В+Мп + Мо (27,9—20,2%) и значительно меньшее после второго полива (14,8%). При внесении бора усиление дыхания через 3 дня после первого полива составляло всего 9,8%, через 6 дней—15,3% и через 3 дня после второго полива — 8%. Небольшое усиление дыхания наблюдалось и при однократном внесении Мп и Мо.
В большинстве случаев повышение интенсивности дыхания при поливе в период созревания значительно ниже, чем при поливе в период цветения и роста. Усиление активности каталазы не совпадает с изменениями интенсивности дыхания по одним и тем же комбинациям.
Большое внимание влиянию некоторых микроэлементов на интенсивность дыхания винограда уделено Р. М. Мехти-Заде (1965) в исследованиях, проведенных в богарных условиях в Азербайджанской ССР.
Оказалось, что действие микроэлементов на интенсивность дыхания проявляется не на всех фазах развития винограда одинаково. Применение микроэлементов перед цветением и в период формирования и роста ягод при 70%-ной влажности почвы от полевой влагоемкости незначительно повышает интенсивность дыхания листьев. Несколько сильнее действует бор и медь при 50%-ной влажности почвы.
В фазу созревания ягод при 70%-ной влажности почвы бор, кобальт и медь снижают активность дыхания по сравнению с контролем. При влажности почвы 50% медь не влияет на интенсивность дыхания, а бор и кобальт несколько снижают ее по сравнению с контролем. Следовательно, для некоторой активизации окислительных процессов целесообразно применение микроудобрений, приуроченное к ранним периодам развития виноградного растения.
Бор, кобальт и медь при различной влажности почвы как при внекорневом, так и корневом питании повышают активность каталазы. Положительное действие бора, кобальта и меди при 50%-ной влажности почвы заметнее, чем при 70%-ной.
По данным Р. М. Мехти-Заде, интенсивность дыхания листьев винограда повышает гиббереллин в концентрации 0,01%. Эффект наблюдается уже через 5 дней после опрыскивания. Параллельно автор наблюдал также усиление йодвосстанавливающей способности тканей листьев (табл. 83).
Таблица 83
Влияние гиббереллина на интенсивность дыхания и йодвосстанавливающую активность листьев
Вариант опыта | Дата опрыскивания | Дата анализа | Интенсивность дыхания, мг СОа на 100 г сырого вещества в 1 ч | Йодвосстанавливающая активность листьев, мг 0.005N | |
8 ч | 13 ч | ||||
Контроль (вода) . | 25/V | 31/V | 41,4 | 48,4 | 702 |
Гиббереллин |
|
|
|
|
|
0,01%-ный | 25/V | 31/V | 48,4 | 52,0 | 825 |
Изучение влияния подвоя на интенсивность дыхания и фотосинтез листьев привоя в связи с выращиванием саженцев на филлоксероустойчивых подвоях показало, что оно довольно значительно (Плакида и др., 1963). Интенсивность фотосинтеза привитых кустов выше, чем корнесобственных (Серексия). Однако подвой Рипария Глуар имеет большее сродство с привоем (Серексия), чем 3309, и интенсивность дыхания у сорта Серексия на последнем подвое очень низкая, возможно из-за более медленного оттока ассимилятов и задержки окислительных процессов. Для сорта Рислинг, очевидно, лучшим будет сочетание с подвоем 3309, чем 1616.
Разработка комплекса физиологических и биохимических показателей для определения лучших пар прививок с большей продуктивностью имеет важное хозяйственное значение. Она позволила бы отказаться от длительных полевых опытов изучения влияния подвоя на привой.
Повреждение винограда хлорозом — частое явление в некоторых районах Грузии, Южной Осетии, Молдавии. Основной причиной, вызывающей хлороз, нужно считать нарушение обмена веществ, ведущее к полному ослаблению растения. Этому наряду с другими причинами может способствовать неправильный подбор прививаемых привоя и подвоя. При сближении двух организмов в прививке могут происходить глубокие биохимические и физиологические изменения в зависимости от степени близости их обмена. По данным Т. Г. Мазанашвили (1953), у сильно хлорозных лоз по сравнению со здоровыми увеличивается интенсивность дыхания, возрастает активность пероксидазы в корнях и листьях, уменьшается титруемая кислотность. При этом идет интенсивная потеря пластических веществ.
В исследованиях Э. Г. Микеладзе (1961) у прививок Ркацители на 3309 по сравнению с корнесобственным Ркацители интенсивность дыхания в большинстве случаев снижается. У привитого Саперави по сравнению с корнесобственным дыхание повышается. Следовательно, у прививок Саперави идет уменьшение накопления сухого вещества, что ведет к ослаблению жизнедеятельности растения. Ясно, что в прививках обоих сортов происходят разные изменения обмена веществ. Одними из первых легкоуловимых изменений обмена являются нарушения окислительно-восстановительных свойств растения. Отсюда наблюдения за такими показателями, как дыхание, активность ферментных систем и др., необходимы при оценке качества прививок и выяснения взаимного влияния подвоя на привой (табл. 84).
Таблица 84
Количество O2, поглощенного листьями (в мл на I г сухого вещества за 1 ч)

На основании своих исследований Э. Г. Макеладзе делает вывод, что привитые растения сильнее повреждаются хлорозом, чем корнесобственные.
При ослаблении жизненности организма происходит десорбция ферментов с повышением при этом их активности (Кружинин, 1955; Чрелашвили, 1951, по Микеладзе, 1961).
Е. А. Чрелашвили (1948) отмечает, что высокой активностью пероксидазы характеризуются не только явно хлороз-ные лозы, но также и те сорта подвойных американских лоз, которые считаются менее хлорозостойкими.
Показатель общей активности окислительных ферментов, как отмечает Э. Г. Микеладзе, под влиянием подвоя у Ркацители снижается, а у Саперави повышается. У корнесобственного Ркацители этот показатель значительно выше, чем у Саперави. Листья подвоя 3309 по этому показателю приближаются к Ркацители, но обладают более выраженной перо-ксидазной активностью (по сравнению с полифенолоксидазной).
Листьям сорта Саперави по сравнению с Ркацители свойственна также большая доля участия пероксидазы в соотношении пероксидазной и полифенолоксидазной активности (табл. 85).
Таблица 85 Активность пероксидазы и полифенолоксидазы (в мл KS03 1/200 N на 1 г сухого вещества за 2 мин)
| Варианты опыта | 1954 г. | 6/VH 1955 г.
| ||||||
общее количество ферментов | общее количество ферментов | пероксидаза | полифенолоксидаза | |||||
5/VI | 20/VII | 30/VII | мл | % от общего количества | мл | % от общего количества | ||
Ркацители корнесобственный | 24,6 | 30,0 | 30,0 | 40,5 | 18,9 | 46,6 | 21,6 | 53,4 |
Ркацители на 3309 . | 16,9 | 22,5 | 30,0 | 31,8 | 14,3 | 44,9 | 17,5 | 55,1 |
Саперави корнесобственный | 14,6 | 9,2 | 19,3 | 30,1 | 18,0 | 59,8 | 12,1 | 40,2 |
Саперави на 3309 | 20,7 | 23,3 | 16,6 | 31,5 | 20,0 | 63,5 | 11,5 | 36,5 |
3309 непривитой | 12,7 | 27,2 | 10,7 | 43,3 | 31,7 | 73,2 | 11,6 | 26,8 |
Прививки способствуют выявлению этого природного свойства Саперави, которое совпадает со свойствами подвоя. У Ркацители прививка усиливает природный сдвиг, наблюдаемый в соотношении деятельности ферментов. Однако этот сдвиг, по результатам автора, не согласуется с соотношением ферментной активности, характерной для подвоя, т. е. под влиянием подвоя у Ркацители возрастает больше не пероксидазная активность, а свойственная ему полифенолоксидазная. Изменения показателей окислительных ферментов в общем аналогичны изменению показателей дыхания (табл. 84).
Автор отмечает, что сдвиг у Саперави не оказывает влияния на качество прививок, но неблагоприятные условия (засуха и др.) могут резко ухудшить их состояние.