Начиная с тридцатых годов нынешнего столетия и в последующее время благодаря исследованиям Р. Зеелингера, Н. П. Кренке, Г. А. Боровикова, А. С. Мержаниана, С. А. Мельника, Л. В. Колесника, А. Г. Мишуренко, Л. М. Малтабара, В. Г. Николенко, И. К. Громаковского и других были разработаны теоретические основы размножения винограда прививкой. Это позволило создать стройную систему выращивания привитых саженцев, которая обеспечивает получение не менее 55—60 % доброкачественных саженцев от числа произведенных прививок.
Анатомия и физиология прививки
Для прививки обычно используют однолетние одревесневшие побеги, разрезанные на черенки различной длины. Известно, что одревесневшие черенки срастаются лишь благодаря образованию каллюса. Первое необходимое условие образования каллюса — наличие раны. Но, как показали исследования Г. А. Боровикова и других, к реакции на раневое раздражение способны не все живые клетки однолетнего побега виноградной лозы. Наибольшей способностью к образованию каллюса обладают клетки камбия, затем клетки коровой паренхимы и сердцевинных лучей и значительно меньше клетки феллодермы и феллогена. Остальные живые элементы вторичной коры непосредственного участия в образовании каллюса не принимают. Кроме наличия раны, необходим и определенный комплекс внешних условий: соответствующая температура, влажность и доступ кислорода воздуха. Скорость нарастания каллюса, а также место и время его появления зависят не только от перечисленных факторов, но и от внутренних условий.
По мнению Кренке, само растение, его состояние внешние условия могут оказывать лишь то или иное влияние на соприкасающиеся клетки подвоя и привоя, главная же роль принадлежит только соприкасающимся клеткам или тканям. Реакция растения на повреждение связана с двумя факторами: с новыми физико-химическими процессами, возникшими непосредственно в области повреждения, с нарушением корреляции частей растений. При прививке в местах разрезов наносят механические повреждения клеткам, из которых вытекает их содержимое. Они претерпевают химические изменения, в результате которых могут появляться различные соединения, отсутствовавшие ранее в живой цитоплазме. Природа этих веществ еще не выяснена.
Присутствием раневых раздражителей можно объяснить, например, явление полярности у виноградной лозы в образовании каллюса. Полярность проявляется в том, что каллюс быстрее и в большом количестве образуется на нижнем конце черенка, позже и в значительно меньших количествах — на верхнем его конце. В связи с тем что при прививке нижний конец привоя соединяется с верхним концом подвоя, образование каллюса происходит раньше и больше на привое и позже и меньше — на подвое. Как показали исследования Г. А. Боровикова, полярность в образовании каллюса на черенках вне прививки выявляется особенно отчетливо, а при прививке энергия образования и масса каллюса на верхнем конце подвоя только немногим уступают его развитию на нижнем конце привоя. Это явление Г. А. Боровиков объясняет диффундированием раневых раздражителей из привоя к подвою, в результате ускоряется процесс образования каллюса у подвоя. Кроме полярности, сильно влияет на его образование дорзивентральность.
Если сделать поперечные разрезы на виноградных черенках и поместить их в благоприятные условия, раньше каллюс образуется на брюшной, а затем на спинной стороне побега (рис. 5). В дальнейшем каллюс разрастается и охватывает сначала плоскую, а затем желобковую стороны. Если побеги хорошо вызрели и сорт отличается хорошим образованием каллюса, смыкание каллюсов брюшной и спинной сторон происходит сравнительно быстро. Если сорт отличается слабым образованием каллюса или черенки недостаточно хорошо вызрели, каллюс на желобковой стороне появляется либо медленно, либо его совсем не бывает.

Рис. 5. Развитие каллюса в зависимости от дорзивентрального строения побега:
1 — каллюс появляется вначале на брюшной стороне, 2 — появление каллюса на спинной стороне. 3 — разрастание каллюса на брюшной и спин, ной сторонах, 4 — каллюс на плоской стороне, 5 — каллюс на желобковой стороне, 6—7 — дальнейшее разрастание каллюса (по Боровикову)
Явление дорзивентральности в образовании каллюса хорошо проявляется только на поперечном (перпендикулярном) срезе. Иное соотношение в скорости появления каллюса наблюдается в том случае, когда на черенках сделаны косые срезы, например при прививке английской копулировкой. В этом случае каллюс развивается прежде всего и лучше там, где копуляционная поверхность образует с наружной поверхностью побега острый угол. Это явление, отмеченное Г. А. Боровиковым, получило название верхушечности (рис. 6). По Бирюкову (Украинской НИИ виноградарства и питомниководства имени В. Е. Таирова), задержка в развитии каллюса у тупого угла вызвана механическими причинами; значительно большим давлением на ткани в момент вхождения режущей части инструмента. Поэтому у тупого угла повреждается больше клеток, чем у острого, в результате каллюс образуется позднее. При прививке обычной косой копулировкой, когда соответствующие стороны подвоя и привоя совпадают и срез проходит так, что язычки находятся на выпуклых сторонах побега, образование каллюса начинается у язычка привоя. Такому явлению способствуют все перечисленные внутренние факторы: полярность, дорзивентральность и верхушечность.

Рис. 6. Развитие каллюса на виноградных черенках в зависимости от явления верхушечности. Вначале каллюс образуется у острого угла на конце среза, и отсюда он постепенно нарастает к тупому углу (по Боровикову)
Значительно позднее каллюс образуется при обычной прививке косой копулировкой на желобковой и плоской сторонах подвоя из-за того, что здесь, кроме отрицательного явления дорзивентральности и верхушечности, действует фактор полярности. Следовательно, направление среза при прививке косой копулировкой имеет большое значение для кругового наплыва каллюса у прививаемых компонентов.
Каллюс сначала образуется не на поверхности раны, а значительно ниже среза, что заметно по утолщению коры. На поверхности срезов подвоя и привоя в результате ранения образуется так называемая изолирующая прослойка, состоящая из мертвых клеток. Изолирующая прослойка бывает особенно толстой и в том случае, если почему-либо задерживается образование каллюса у одного из прививаемых компонентов. Для создания наиболее благоприятных условий одновременного и кругового наплыва каллюса по всей окружности копуляционных срезов подвоя и привоя необходимо:
а) свести до минимума отрицательное явление полярности в образовании каллюса. Этого достигают либо предпрививочным вымачиванием подвоя при повышенной температуре, либо воздействием стимуляторов, либо предпрививочной стратификацией, а также более плотным соединением прививаемых компонентов для лучшего диффундирования раневых веществ из привоя в подвой;
б) устранить хотя бы частично отрицательное явление дорзивентральности, чего можно достигнуть изменением направления среза на подвое. Срез необходимо делать так, чтобы тупой угол приходился на брюшную или спинную (наиболее развитые) стороны побега. При таком направлении среза язычки для соединения прививаемых компонентов зарезают на желобковой и плоской сторонах, наименее обеспеченных питательными веществами из-за дорзивентрального строения побега. У них процесс образования каллюса будет немного ускоряться благодаря положительному действию явления верхушечности. Это же явление будет немного задерживать образование каллюса на самой развитой (брюшной) стороне побега. В результате каллюс будет образовываться более равномерно по всей окружности среза.
Исследования Н. П. Кренке, Г. А. Боровикова и других показали, что в образовании изолирующей прослойки принимают участие мертвые, перерезанные ножом клетки коры, камбия, отмершие, непосредственно прилегающие к первым, перераздраженные и наружные клетки каллюса со слегка опробковевшими оболочками.
Толщина изолирующей прослойки зависит от степени сродства между привоем и подвоем, условий, в которые попадают прививки, и энергии образования каллюса на подвое и привое. Чем раньше после прививки начинается образование каллюса в местах повреждения, тем меньше будет слой мертвых клеток и, следовательно, тоньше изолирующая прослойка. Если каллюс образуется одновременно по всей окружности среза на подвое и привое, наплывы каллюса обоих прививаемых компонентов соприкасаются быстрее. Оболочки наружных клеток каллюса в этом случае меньше пробковеют, и, следовательно, тоньше будет изолирующая прослойка. И, наоборот, если образование каллюса у одного из компонентов или на одной из сторон черенка начинается раньше, чем у другого компонента, или на другой стороне черенка, то на поверхности этих ранее образовавшихся наплывов каллюса верхний слой клеток пробковеет больше и изолирующая прослойка получается толще. Толщина прослойки может значительно изменяться также в зависимости от качества и способов прививки и прививаемых черенков, и от характера и напряженности ферментативных процессов (Л. В. Колесник).
Наиболее полные исследования по анатомии срастания у винограда осуществлены Г. А. Боровиковым.
При благоприятных условиях внешней среды у прививаемых компонентов под влиянием ранения клетки камбия и коры начинают энергично делиться и образуют наплывы каллюса. Клетки каллюса вследствие взаимного сжатия имеют неправильную форму и различную величину, с очень тонкими и нежными целлюлозными оболочками и плотно соединены друг с другом. На поверхности каллюса вначале образуется сплошной слой опробковевших клеток изолирующей прослойки, который в дальнейшем при энергичном делении клеток каллюса разрывается на отдельные участки разной величины в зависимости от плотности мертвых клеток. Образуются так называемые окна прорыва, через которые происходит спайка каллюсов прививаемых компонентов.
Изолирующая прослойка особенно резко выступает (что представляет наибольшие трудности для спайки) в том случае, когда каллюс одного из компонентов образовался раньше другого или каллюс одного из компонентов почему-либо приостановил свое развитие. Если образующиеся каллюсы обоих компонентов соприкасаются быстро, т. е. в период энергичного деления их клеток, то изолирующая прослойка не образует сплошной линии, а заметна лишь в виде небольших островков по линии спайки. В тех случаях, когда на большом протяжении оба каллюса соприкасаются «окнами прорыва», спайка клеток каллюса обоих компонентов происходит настолько прочно, что порой нельзя установить, где кончаются клетки каллюса одного компонента и начинаются клетки другого.
Клетки каллюса одного из компонентов могут также хорошо спаиваться с клетками изолирующей прослойки другого компонента. Это соединение бывает настолько плотным, что при попытке механического разъединения таких участков разрываются живые ткани, а живые клетки каллюса не отделяются от мертвых клеток изолирующего слоя. Благодаря такой прочной спайке живых клеток с мертвыми, по мнению Г. А. Боровикова, и рассасывается изолирующая прослойка. Рассасывание прослойки происходит тем легче, чем она тоньше. Часто сравнительно тонкая изолирующая прослойка рассасывается полностью, но нередко, когда в состав ее входило много мертвых клеток, полного рассасывания не наблюдается.
В тех случаях, когда участки изолирующей прослойки лежат не на пути вновь образованных камбиальных слоев, отрицательного значения нерассосавшиеся участки изолирующей прослойки не имеют. Когда участки изолирующей прослойки лежат на пути следования вновь образующегося камбия, они могут задержать или даже воспрепятствовать срастанию при. вивок в данном месте. В этом случае камбий, дойдя до изолирующего участка, направляется от него в сторону. Нередки случаи, когда камбий привоя и подвоя направляется в противоположные стороны, что задерживает или даже полностью устраняет возможность сращения. Под сращением привоя с подвоем обычно понимают такое состояние, при котором в результате спайки камбия привоя и подвоя устанавливается связь между проводящими элементами. Камбий в каллюсах привоя и подвоя образуется значительно раньше их спайки. Г. А. Боровиков неоднократно наблюдал образование камбия в различных точках каллюса даже у таких прививок, где не было соприкосновения каллюсов прививаемых компонентов.
В дальнейшем для успешного срастания образовавшегося камбия важно, чтобы на его пути не было препятствий, которые изменяли бы его желаемое направление. Одним из таких препятствий нередко бывает толстая изолирующая прослойка, образующаяся при длительной консервации прививок, и потоки каллюса при длительной стратификации. Г. А. Боровиков наблюдал, что иногда еще задолго до срастания камбия проходит его дифференциация и образуются элементы ксилемы и флоэмы, причем ксилема всегда образуется раньше флоэмы. Образование элементов ксилемы, по наблюдениям Л. В. Колесника, на различных сторонах побега идет неравномерно: они появляются прежде всего там, где раньше образовался каллюс.
Элементы ксилемы (сосуды), которые должны соединить друг с другом соответствующие элементы подвоя и привоя, переходят линию спайки не по прямому направлению, а загибаясь и образуя прямой угол или колено. Эти сосуды всегда очень короткие и извилистые. В результате скорее и лучше устанавливается связь между проводящими элементами. Вскоре после появления промежуточного камбия, который соединяет привой с подвоем и образует элементы ксилемы и флоэмы, в последней образуется пробковый камбий— феллоген, который соединяется с феллогеном привоя ,и подвоя. В результате деятельности феллогена образуется перидерма, которая отделяет лишние наружные наплывы каллюса подвоя и привоя, и они отбирают.
Большая часть внутренних наплывов каллюса также пробковеет и отмирает, но здесь значительное число клеток каллюса остаются живыми, особенно те, которые расположены непосредственно под древесиной и появились в результате деятельности новообразованного камбия. В этом месте оболочки клеток древеснеют, но остаются живыми и являются местом отложения крахмала. В местах прививки, где не произошло срастания подвоя с привоем, наблюдаются местные гипертрофии, достигающие на 2—3-й год больших размеров. Это наблюдается также при неудачном подборе комбинаций привоя с подвоем (даже в случае их хорошего срастания) как результат задержки передвижения питательных веществ.
Для успешного срастания прививаемых компонентов большое значение имеет укоренение подвоя. Иногда низкий выход привитых саженцев из школки объясняется не столько плохим образованием каллюса по окружности копуляционного среза, сколько плохим укоренением растений в школке. Во время стратификации и обычной, применяемой в производстве, закалки привитых черенков в специальных помещениях или парниках наблюдается лишь частичная спайка каллюсов прививаемых компонентов. Окончательное сращение прививок с образованием сосудистой системы в результате деятельности промежуточного камбия происходит в естественных условиях в школке. Чем быстрее укореняются прививки после посадки в школку, тем больше выход саженцев с круговым срастанием.
Объясняется это тем, что в открытом грунте трудно создать нормальные условия увлажнения у места спайки, поэтому часто наплывы каллюса подсыхают. К моменту посадки привитых черенков в школку они должны быть не только спаяны наплывами каллюса, но и хорошо подготовлены к быстрому укоренению.
На побегах виноградной лозы нет корневых зачатков, как у многих других растений (ива, черная смородина и др.); они закладываются лишь после того, как черенки попадают во влажную среду при повышенной температуре и хорошем доступе воздуха. У первой исходной клетки корневого зачатка движущим началом ее саморазвития в сторону образования нового органа корневого типа должны быть те же функции поглощения и усвоения веществ, которые присущи корням.
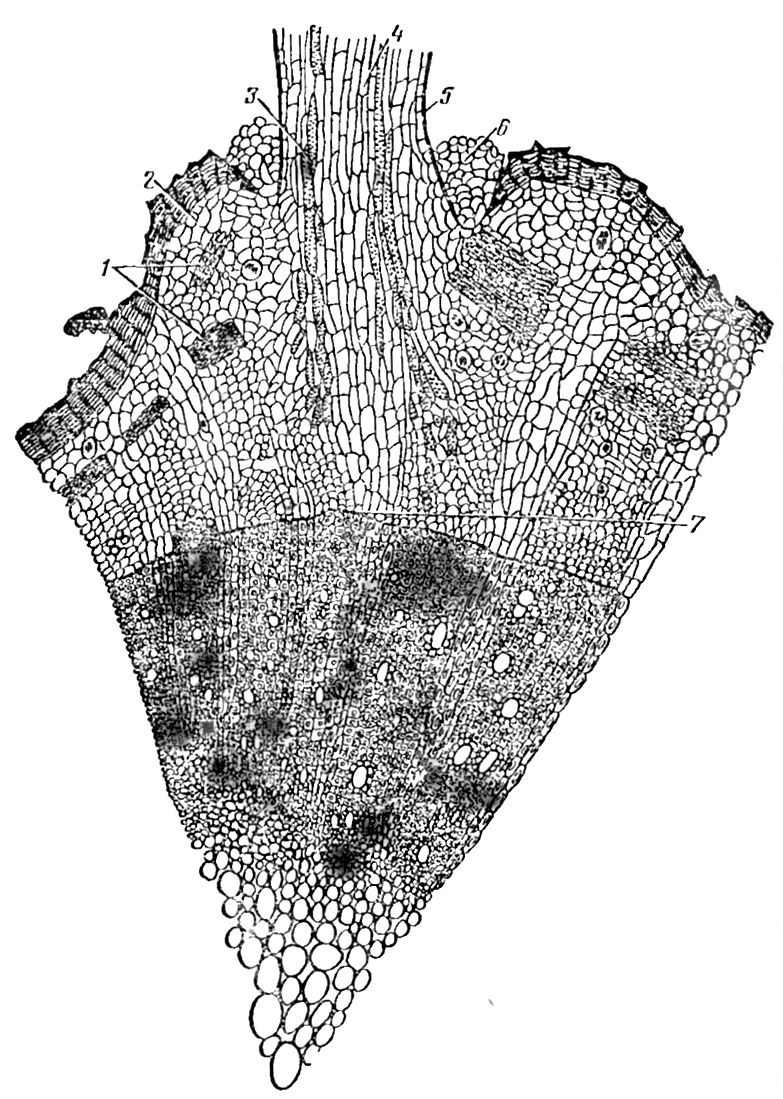
Рис. 7. Закладка придаточного корня у черенка:
1 — группа лубяных волокон, 2 — перидерма, 3 — сосуды корешка, 4 - сердцевина корешка, 5 — кора корешка, 6 — остатки «кармашка» у основания корешка, 7 — первичный луч, над которым заложился корешок (по Баранову)
Отличие состоит в том, что сформировавшийся корень извлекает воду и растворенные вещества из почвы. Молодой корневой зачаток, считая от первой инициальной клетки, поглощает необходимые воду и питательные вещества из окружающих взрослых клеток материнского органа. В связи с тем что к основанию черенка прилегает сравнительно влажная среда (почва, влажные опилки, мох и др.), рост корня, согласно гидротропизму, направлен к периферии черенка.
У виноградных лоз адвентивные корни закладываются в периферийной зоне первичных сердцевинных лучей, которые возникли из перицикла. Клетки перицикла, прилегающие непосредственно к периферийному слою клеток сердцевинного луча, начинают делиться и образуют бугорок меристематической ткани в виде конического тельца. Вершина бугорка обращена к периферии черенка. Одновременно с формированием зачатка корня пробуждается деятельность и в окружающих его тканях сердцевинного луча, а также в прилегающих участках флоэмы. В результате деления клеток образуется рыхлая ткань с тонкостенными клеточными оболочками. Эта рыхлая ткань разрастается и раздвигает в стороны флоэмные участки, смыкается над зачатком корешка и давит на перидерму. К этому времени растущий зачаток корешка уже имеет на кончике корневой чехлик, которым давит на эту рыхлую ткань, называемую кармашком корешка. В результате перидерма разрывается, и в этом месте сначала появляется белый бугорок рыхлой ткани, а затем уже корешок прорывает бугорок и выходит наружу (рис. 7).
Следовательно, для хорошего и быстрого укоренения в почве прививки должны быть до посадки подготовлены так, чтобы у места соединения подвоя и привоя была прочная спайка каллюсов, а у основания подвоя образовались зачатки корешков длиной 2—3 мм.