Перекисная теория, сформулированная А. Н. Бахом в 1897 г., в сущности была первой теорией биологического окисления. Согласно этой теории механизм биологического окисления рассматривается следующим образом: при активации в молекуле кислорода разрывается одна из связей, удерживающих атомы. В результате образуется перекисная группа —О—О—, которая присоединяется к окисляющемуся соединению, образуя перекись.
При воздействии на эту перекись фермента пероксидазы окислительный потенциал значительно увеличивается вследствие отщепления от перекиси активного кислорода, который способен вызвать окисление более трудноокисляемого соединения.
А. Н. Бах объяснял механизм биологического окисления с позиции теории перекисного окисления, согласно которой молекулярный кислород присоединяется к ненасыщенному, легкоокисляемому веществу, названному оксигеназой. Механизм окисления с помощью оксигеназы и пероксидазы показан ниже:
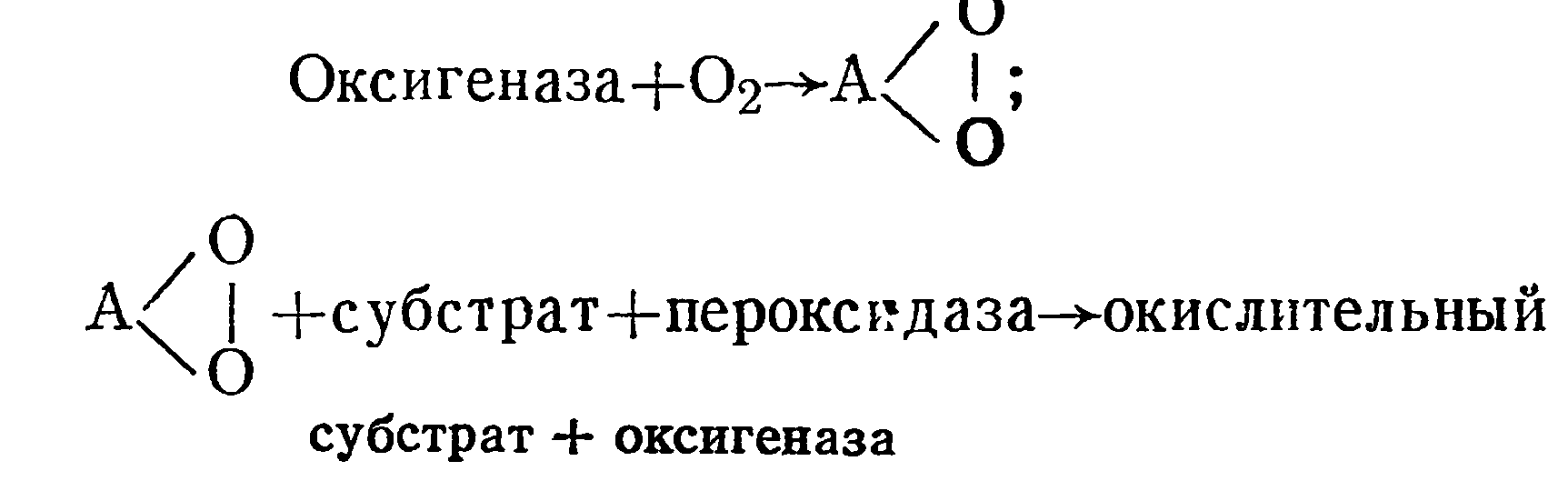
Не так давно О. Хаяши (1964) и Г. Мезон (1957) сообщили, что в растениях и микроорганизмах содержится фермент типа оксигеназ, который активирует кислород воздуха, делая его способным вклиниваться в окисляемые вещества.
Мезон назвал этот фермент «трансферазой кислорода», а О. Хаяши предложил баховский термин «оксигеназы». Существуют высокомолекулярные ферменты, так называемые монооксигеназы и диоксигеназы.
Открытие нового фермента «оксигеназы», активирующего молекулярный кислород и прямо внедряющегося в субстрат, подтвердило правильность переписной теории биологического окисления А. Н. Баха.
За последнее время особое значение придают супероксиду [иону] в окислительно-восстановительных процессах [120], является высокоактивным ионным радикалом, который образуется при одноэлектронном восстановлении молекулярного кислорода, а также при энзиматическом аэробном метаболизме. Этот супероксид обладает как окислительными, так и восстановительными свойствами в зависимости от условий проведения реакции (pH среды). Это не противоречит правильности перекисной теории А. Н. Баха, так как в данном случае мы также имеем активирование кислорода.
Однако наряду с теорией перекисного окисления, которая предусматривает активирование кислорода, была выдвинута теория дегидрирования, т. е. отнятия водорода в процессе биологического окисления. Впервые теорию дегидрирования выдвинул В. И. Палладии (1912), который считал, что для завершения биологического окисления наряду с активированием кислорода необходимо и активирование водорода субстрата.
Г. Виланд подтвердил более раннее представление В. И. Палладина, который считал, что в процессе биологического окисления главная роль принадлежит лабилизации водорода органического вещества, переходящего затем в акцептор кислорода, дающий с водородом воду.
Согласно теории Г. Виланда, специфичность фермента проявляется только лишь по отношению донатора водорода, акцепторная специфичность им отрицалась.
В. И. Палладии впервые показал, что дыхание в растениях осуществляется при помощи особых посредников дыхания. К посредникам относятся полифенолы, которые окисляются кислородом под действием оксидаз, приобретая ту или иную окраску и превращаются в пигменты, так называемые хромогены.
Этот исследователь также показал, что хромоген активирует не только кислород, а также водород дыхательного субстрата в присутствии дегидрогеназ. В. И. Палладии заменил дыхательные хромогены на водородные акцепторы. Он объединил процесс окисления и восстановления при непосредственном участии воды. Так, например, водород воды восстанавливает дыхательный пигмент и превращает его в хромоген, а кислород воды окисляет хромоген в пигмент. В. И. Палладии придавал особое значение участию воды в процессе дыхания, считая, что в образовании СO2 участвует кислород воды, а не кислород атмосферы. Это было доказано экспериментально с помощью изотопа кислорода Б. Б. Вартапетяном и А. Л. Курсановым (1955).
Теория В. И. Палладина хорошо объясняет жизнь анаэробных микроорганизмов, которые в своей жизнедеятельности обходятся без кислорода. В этом случае акцептором водорода при дегидрировании органического вещества является не кислород воздуха, и следовательно, вода не образуется. Акцептором водорода при алкогольном брожении может служить уксусный альдегид, который восстанавливается в этанол, а при молочнокислом брожении акцептором водорода является пировиноградная кислота, которая также восстанавливается в молочную кислоту. В ацетобутиловом брожении акцептором водорода является масляный альдегид, который восстанавливается в бутанол.
Теория окислительно-восстановительных процессов - Перекисная теория окисления вин
Содержание материала
Страница 2 из 5
- Вы здесь:
- Главная

- Статьи
- Книги
- Основы биохимии виноделия
- Теория окислительно-восстановительных процессов