Установление синтетических функций корневой системы растения в отношении алкалоидов, аминокислот и других физиологически активных веществ явилось стимулом в поисках синтетических проявлений корневой системы виноградной лозы. Усилия физиологов в области виноградарства были направлены на установление синтеза аминокислот в корнях лозы. С т о е в с сотр. (1959, 1959а) проводили хроматографический анализ сахаров и свободных аминокислот пасоки сорта Гымза, выращенного без прививки и с прививкой на подвой Рупестрис дю Ло. Анализировалась также пасока подвойной лозы Рупестрис дю Ло. Несколько позже Дурмишидзе и Хачидзе (1959, 1960) изучали органические кислоты и содержание аминокислот в пасоке виноградной лозы сортов Саперави и Ркацители, подвоя Берландиери х Рипария Кобера 5ББ и гибридов Бета и Оберлен. Основные выводы, вытекающие из исследований синтетических функций корневой системы винограда обеих коллективов, сводятся к следующему.
- Аминокислотный состав пасоки виноградной лозы одинаков в различных географических районах.
При исследовании в Болгарии и Грузинской ССР в пасоке установлены аспарагиновая и глютаминовая кислоты, валин, изолейцин, лизин, аланин, тирозин, фенилаланин, гистидин, цистин, пролин, аргинин, гликокол, метионин, γ-аминомасляная кислота и глютамин. Существенных различий в аминокислотном составе пасоки в этих столь различных географических районах не обнаружено.
- Набор аминокислот изменяется по фазам вегетации.
По данным Стоева с сотр. (1959, 1959а), в начале сокодвижения в пасоке виноградной лозы содержалось ограниченное число аминокислот: аспарагиновой и глютаминовой кислот, валина и изолейцина. В последующие сроки сбора пасоки количество аминокислот в ней возрастало: 10 апреля в пасоке было обнаружено 9 аминокислот. В ней появились лизин, аланин, тирозин, фенилаланин и одно неидентифицированное вещество. В третий срок (14/V) наблюдалось еще большее увеличение аминокислотного набора пасоки. Помимо перечисленных выше аминокислот, в ней были обнаружены гистидин, цистин, а также несколько неидентифицированных веществ.
В пасоке, полученной в начале цветения (5/VI), отмечалось уменьшение набора аминокислот. Их число снижалось до 7—9. В это время исчезали цистин и тирозин, однако появлялись пролин и аланин. К концу июня и в начале июля набор аминокислот в пасоке еще больше уменьшался; установлено лишь 4 аминокислоты: лизин, аспарагиновая и глютаминовая кислоты и пролин.
Результаты этих исследований показали, что при передвижении аминокислот из корневой системы к надземным органам их качественный состав не изменялся. Он не изменялся также при переходе через спайки прививки.

Рис. 7. Сезонные изменения аминокислот в пасоке виноградной лозы
а — начало сокодвижения; б — разгар сокодвижения; в — длина побегов 40—50 см; г — разгар цветения; 1 — лейцин; 2 — фенилаланин; 3 — валин; 5 — тирозин; 6 — пролин; 7 — аланин; 8 — треонин; 9 — глютаминовая кислота; 10— серин+глицин; 11 — глютамин; 12 — аспарагин; 13 — лизин; 14 — аспарагиновая кислота; Е — экстинкция

Рис. 8. Суточные изменения аминокислот (26/IV—27/IV 1963 г.)
а — 22—04 ч.; б — 04—10 ч.; в — 10—16 ч.; г — 16—22 ч.; 1 — лейцин; 2 — фенилаланин; 3 — валин; 6 — пролин; 7 — аланин; 8 — треонин; 9 — глютаминовая кислота; 10 — серин+глицин; 11 — глютамин; 12 — аспарагин; 13 — лизин; 14 — аспарагиновая кислота; Б — экстинкция
Дальнейшее исследование сезонных изменений аминокислот в пасоке виноградной лозы (Стоев с сотр., 1965; Stoev c сотр., 1966) показало, что аминокислотный набор увеличивается главным образом в разгаре сокодвижения и в период цветения (рис. 7). Из идентифицированных веществ в наибольшем количестве находятся глютамин, глютаминовая кислота и аланин.
В начале сокодвижения значительное место занимают аспарагин и лизин, а позже валин и пролин.
Дурмишидзе и Хачидзе (1960) также установили, что состав аминокислот в пасоке значительно изменяется в течение вегетации — как число свободных аминокислот, так и их общее количество наиболее велики в период сокодвижения.
- Различий в составе аминокислот пасоки сортов не имеется.
К такому выводу пришли Стоев с сотр. (1959, 1959а), Былик и Стоев (1975), Дурмишидзе и Хачидзе (1959, 1960). Коzmа и сотр. (1963) сообщает о сортовых различиях в аминокислотном составе пасоки.
- Набор аминокислот изменяется в течение суток.
Изучение суточных изменений аминокислот в пасоке виноградной лозы (Stоеv с сотр., 1966) показало, что синтез аминокислот неодинаков в различные часы суток (рис. 8). Наибольший набор аминокислот имеется в пасоке, собранной в интервале с 4 до 10 ч. утра. Количественное содержание аминокислот также наиболее высоко в ранние утренние часы. Большой набор и большое количество аминокислот наблюдаются в дневные часы — с 10 до 16 ч. Набор аминокислот в послеполуденные часы уменьшается и достигает минимума в 22 ч. Ночью (22—04 ч.) отмечается некоторое увеличение числа аминокислот.
Из идентифицированных веществ в наибольшем количестве встречаются глютамин, глютаминовая кислота, аланин, пролин, аспарагин и валин. Из сахаров в пасоке обнаружены только фруктоза и глюкоза, которые выявлялись все время на протяжении суток (рис. 9).
Мокроносов с сотр. (1957) также установили суточные изменения в качественном составе аминокислот пасоки картофеля. Между 10 и 17 ч. набор аминокислот в пасоке сильно уменьшался. Впоследствии между 22 и 3 ч. ночи число аминокислот продолжало уменьшаться.
Количественный анализ также выявил суточную ритмичность в содержании сахаров и аминного азота (Стоев с сотр., 1959, 1959а). Максимум выделения сахаров и аминокислот из корней достигался в 8—14 ч., после чего их количество постепенно уменьшалось и было минимальным в 20 ч. Ночью содержание сахаров и аминного азота в пасоке снова увеличивалось и к 8 ч. утра приближалось к максимальной величине.

Рис. 9. Суточная динамика содержания сахаров и аминного азота в пасоке виноградной лозы сорта Зарчин (привитого) (22—23/IV 1957 г.)
а — сахара; б — аминный азот
- Набор аминокислот изменяется под влиянием минерального удобрения.
С целью установить, вызывают ли минеральные вещества при их поглощении корнями изменения в синтезе аминокислот, проведен ряд исследований. Уже первые исследования (Стоев с сотр., 1959, 1959а) показали, что аммиачный азот переходит в корнях виноградной лозы в органическую форму и оттуда в виде различных аминокислот передвигается в восходящем направлении. При этом наиболее интенсивное превращение органического азота в аминокислоту происходит в первые дни после его внесения в почву. Авторам удалось установить различия в аминокислотном составе в пасоке удобренных и неудобренных кустов на второй день после внесения удобрения.

Рис. 10. Изменение аминокислотного состава пасоки виноградной лозы под влиянием удобрения азотом (перед цветением; 8/V 1958 г.)
а — на 1-й день; б — на 2-й день; в — на 5-й день; г — на 10-й день; д — на 15-й день; 1 — аспарагиновая кислота; 2 — глютаминовая кислота; 3 — гликокол; 4 — серин; 5 — треонин; б — аспарагин; 7 — глютамин; 8 — аланин; 9 — тирозин; 10 — валин; 11 — лейцин+изолейцин; 0 — неидентифицированные вещества
Дальнейшее изучение влияния минерального удобрения на синтез аминокислот в корневой системе виноградной лозы (Стоев с сотр., 1965; Stоеv с сотр., 1966) показало, что в пасоке до 2—5-го дня после внесения удобрения количество аминокислот наиболее велико, после чего начинает уменьшаться. Особенно резко уменьшается набор аминокислот после десятого дня.
На рис. 10—13 представлены хроматограммы аминокислот в пасоке виноградных кустов, удобренных N, NK, NP, NPK (хроматограммы аминокислот остальных вариантов вполне аналогичны)*.
* Удобрение производилось N, Р, К, NP, РК, NK и NPK, а определение аминокислот — методом хроматографии на бумаге через 1, 2, 5, 10 и 15 дней после внесения удобрений.
В первый день после внесения удобрения аминокислот (в том числе неидентифицированных веществ) больше десяти — в среднем 12,1 (по всем вариантам). На 2—10-й день среднее число идентифицированных и неидентифицированных веществ — порядка 8—9, а на 15-й снижается в среднем до 5—7.

Рис. 11. Изменение аминокислотного состава пасоки виноградной лозы под влиянием удобрения NK (перед цветением; 8/V 1958 г.)
а — д, 1—11, 0 — как на рис. 10
Содержание аминокислот относительно наиболее высоко на 1-й, 2-й и 5-й день после внесения удобрения. Это дает основание предполагать, что поглощение минеральных веществ и их включение в обмен веществ в корневой системе протекает с наибольшей интенсивностью в первые дни после внесения удобрений в почву. После десятого дня как будто наступает затухание в поглощении и превращении минеральных веществ. Об этом свидетельствуют данные табл. 3 о содержании аминного и общего азота в пасоке виноградной лозы.
Полное подтверждение этот вывод нашел значительно позже в исследованиях Нуцубидзе (1974). Подытоживая свои исследования, автор пишет: "Включение азота в свободные аминокислоты корней и листьев происходит в первые же минуты. Максимальная интенсивность поглощения — ассимиляция азота виноградной лозой — падает на первые два дня от начала подкормки" (с. 235).
Аналогичные выводы вытекают и из работы Petrie и Wood (1938). По их исследованиям, на следующий день после обильной азотной подкормки в листьях райграса устанавливается значительное повышение количества амидов, аминокислот и белков.

Рис. 12. Изменение аминокислотного состава пасоки виноградной лозы под влиянием удобрения NP (перед цветением; 8/V 1958 г.)
а — д, 1—11, О — как на рис. 10
Содержание общего и аминного азота в пасоке виноградной лозы (в γ на 1 см жидкости) (9/V—23/V 1958 г.)
Таблица 3
| Варианты | Аминный азот | Общий азот | ||||||||
1-й | 2-й | 5-й | 10-й | 15-й | 1-й | 2-й | 5-й | 10-й | 15-Й | |
Водный контроль | 25 | 39 | 20 | 14 | 14 |
| 119 | 70 | 60 | 38 |
Сухой контроль | 28 | 56 | 28 | 14 | 14 | 238 | 204 | 96 | 107 | 100 |
N | 36 | 112 | 28 | 28 | 28 | 268 | 159 | 127 | 169 | 79 |
Р | 28 | 50 | 34 | 14 | 14 | 267 | 193 | 116 | 104 | 41 |
К | 48 | 78 | 20 | 17 | 14 | — | 166 | 90 | 169 | 54 |
NP | 34 | 64 | 28 | 28 | 14 | 266 | 221 | 86 | 139 | 38 |
NK | 42 | 56 | 39 | 17 | 14 | 281 | 258 | 154 | 153 | 38 |
РК | 36 | 56 | 39 | 17 | 14 | 260 | 222 | 97 | 75 | 38 |
NPK | 34 | 48 | 28 | 20 | 14 | — | 168 | 120 | 100 | 72 |
На второй день после подкормки обнаруживается лишь незначительное увеличение содержания белков и амидов, в то время как содержание небелкового органического азота все еще продолжает сильно увеличиваться. На третий день после подкормки в содержании как белкового, так и небелкового азота уже никаких изменений не отмечалось.
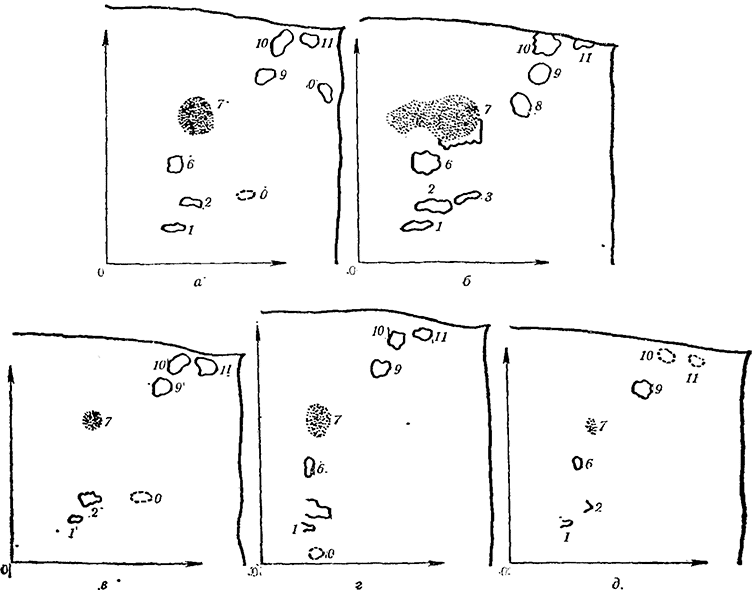
Рис. 13. Изменение аминокислотного состава пасоки виноградной лозы под влиянием удобрения NPK (перед цветением; 8/V 1958 г.)
— д, 1—11, О — как на рис. 10 а
По данным Кретовича (1958), Кретовича с сотр. (1957, 1959, 1959а), Warburg c сотр. (1957), Suzuki с сотр. (1958), глютаминовая кислота является одним из наиболее подвижных метаболитов в растении, играющим очень важную роль в обмене веществ. Глютаминовая кислота быстрее других аминокислот подвергается окислению, синтезу аммиака и кетокислот (Кретович, Яковлева, 1957), быстрее включается также в реакцию переаминирования. Она является источником аминных групп, которые используются для синтеза аспарагина и глютамина. По мнению Плешкова с сотр. (1959), роль в прогрессивном метаморфозе азота и обмена аминокислот принадлежит глютаминовой кислоте и продуктам ее превращения.
Дальнейшие исследования Durmischidze (1964) дают представление об обмене глютаминовой кислоты в виноградном растении. Они показывают прежде всего, что 60 % введенной глютаминовой кислоты — 514С в стеблевых частях и корнях винограда в течение 30 min — превращается в другие аминокислоты, в органические кислоты и частично в сахар (только в побегах). Согласно полученным данным, радиоактивный углерод глютаминовой кислоты включается главным образом в образование лейцина, метионина, фенилаламина, яблочной кислоты и сахарозы (только в побегах).
Несмотря на большую подвижность глютаминовой кислоты, глютамину, по-видимому, принадлежит немаловажная роль в обмене азотистых веществ в корневой системе виноградной лозы. Об этом свидетельствует хотя бы то обстоятельство, что глютамин превалировал по своему количественному содержанию в пасоке виноградной лозы во всех проведенных Стоевым с сотр. опытах и на всех этапах исследования (1959, 1959а, 1960, 1965; Stoev c сотр., 1966). Достаточно взглянуть на хроматограммы, представленные на рисунках, и их дензитометрическое измерение, чтобы это не вызывало ни малейшего сомнения.
Этот факт полностью подтвердился при культуре томатов, где с помощью меченых атомов установлено, что глютамин занимает одно из центральных мест в метаболизме корней. Нуцубидзе (1974, 1977) выражает мнение, что ассимиляция азота происходит путем преимущественного его включения в состав свободных аминокислот, а не амидов. Среди них интенсивно образуются глютаминовая и аспарагиновая кислоты, L-аланин, аргинин и тирозин.
По данным Кретовича и Яковлевой (1957), при ассимиляции аммиака растениями и включении его в различные органические азотистые соединения первостепенная роль принадлежит не только глютаминовой кислоте, синтезируемой из L-кетоглютарата и аммония под действием глютаминдегидразы, но также глютамину, чрезвычайно интенсивно синтезируемому из глютаминовой кислоты и аммония при участии глютаминсинтетазы. По их мнению, глютамин используется в реакциях переаминирования с кетокислотами и может подвергаться дезаминированию под воздействием глютаминазы. Чрезвычайно важно, что глютамин играет роль главнейшего источника аммиачных групп, потребляемых при реакции переаминирования с кетокислотами.
На отдельных этапах сокодвижения аланин также появляется в большом количестве, и считается несомненным, что в процессе ассимиляции аммиачных солей растительным организмом L-аланину принадлежит важная роль.
Вопрос о метаболизме аланина в побегах и корнях виноградной лозы в значительной степени выяснен исследованиями Durmischidze (1964).
При введении аланина 1-14С в ткани растения радиоактивный углерод переходит в другие аминокислоты, сахара (преимущественно в побегах) и органические кислоты (преимущественно в корнях). Как показывают результаты Дурмишидзе, аланин 1-14С переходит в радиоактивную аспарагиновую кислоту, глютаминовую кислоту, пролин, фенилаланин и метионин. В корнях радиоактивность метионина превышает треть радиоактивности всех аминокислот. Радиоактивный углерод аланина включался также в глюкозу, фруктозу, янтарную и гликолевую кислоты (только в побегах).
Следует остановиться и на γ-аминомасляной кислоте. Наличие ее в пасоке виноградной лозы было установлено при изучении влияния удобрения, а также прц изучении суточных и сезонных изменений аминокислотного состава, что свидетельствует об определенной роли этой аминокислоты в обмене азотистых веществ в корнях лозы.
Дальнейшее исследование синтетических функций корневой системы проводилось с использованием меченого источника азота (Dintscheff с сотр., 1964). Целью этого исследования являлось установление времени поступления азота в корни виноградной лозы и его метаболизма в корневой системе. Удобрение сернокислым аммонием проводилось в разгаре сокодвижения сорта Болгар, привитого на подвой Рупестрис дю Ло. Сбор пасоки производили через 4, 8 и 12 h и через 2, 3, 5, 8, 12 и 27 дней после внесения удобрения.
Результаты исследования, приведенные в табл. 4, показывают, что основной формой азота в пасоке винограда, выделенной во время сокодвижения и начала роста, является органическая. На втором месте стоит нитратная форма азота и меньше всего — аммиачного азота.
Таблица 4
Химический и изотопный анализ различных форм азота в пасоке виноградной лозы

Стабильный изотоп найден и во всех трех фракциях — аммиачной, нитратной и органической. Больше всего 16N содержится в нитратной фракции, меньше — в аммиачной. Содержание 15Ν в органической фракции занимает промежуточное место — 6,5—55,09 mg/dm3 пасоки.
Результаты показывают также, что азот начинает поступать в корневую систему лишь через три дня после внесения удобрения. Более медленное поступление азота, чем было установлено раньше (Стоев с сотр., 1959, 1959а), вероятно, обусловлено понижением температуры, отмечавшимся после внесения удобрения.
Наличие органического азота в пасоке, представленного стабильным изотопом (l6N), указывает, что в корнях виноградной лозы проходят синтетические процессы и что корневая система обладает способностью синтезировать аминокислоты и амиды.
Нуцубидзе (1974) в своем исследовании, посвященном проблеме ассимиляции азота виноградной лозой, полностью подтверждает сделанные нами основные выводы (Стоев с сотр., 1959, 1959а, 1960,1965; Стоев с сотр., 1966; Dintscheff с сотр., 1964), сформулированные в отдельных работах и в монографии "Физиологические основы виноградарства" (Стоев, 1971). Вместе с тем он значительно обогащает знания в области ассимиляции азота виноградной лозой и его метаболизма. Автор устанавливает прежде всего, что ассимиляция азота виноградной лозой в основном происходит так же, как у однолетних растений (Кретович и др., 1965, 1967; Меistеr, 1968). Виноградная лоза, отмечает Нуцубидзе (1974), при образовании аминокислот из поглощенного растением азота использует различные кетокислоты ЦТК (цикла трикарбоновых кислот), обеспечив в конечном итоге необходимый набор аминокислот. Если по какой-либо причине цикл Кребса перекрывается, то образование основных кислот, акцепторов аммиака не прекращается, так как открываются другие пути метаболизма — глиоксилатный, фотосинтетический циклы. Не исключаются также взаимопревращения кетокислот вне перечисленных циклов.
Важное значение имеет заключение автора, что в виноградной лозе при ассимиляции азота накопляются главным образом свободные аминокислоты, среди которых преобладают аспарагиновая и глютаминовая кислоты, аланин, аргинин, тирозин и др., тогда как в однолетних растениях происходит преимущественное накопление амидов. Этот факт, по мнению Нуцубидзе, указывает на специфическую направленность азотного обмена виноградной лозы и его саморегулирование.
О синтетических функциях корневой системы виноградной лозы свидетельствует также наличие ряда других веществ в выделенной корнями пасоке. По сообщениям Чкуаселии Тарасашвили (1959), в пасоке имеются аневрин и рибофлавин, причем их содержание подвержено значительным изменениям в ходе сокодвижения лозы в течение суток. Позже Чкуасели (1977) указывает, что условия минерального питания влияют на выделение витаминов корнями. Формы азота влияют на выделение тиамина и рибофлавина. В корнях он обнаружил также хлорофилл а и б, каротин и неоксантин. Установлена связь между синтезом пигментов в корнях и превращением СO2 в корневой системе.
Хачидзе (1957) обнаружил в пасоке виноградной лозы витамины группы В, а Дурмишидзе (1955) — таниновые вещества.
Дурмишидзе и Хачидзе (1959, 1960) установили в пасоке винограда пировиноградную, L-кетоглютаровую, щавелевоуксусную, яблочную, лимонную, янтарную и фумаровую кислоты. По количеству, однако, преобладает яблочная кислота. По мнению авторов, кислоты ди- и трикарбонового цикла и аминокислоты переходят из корней в надземные органы восходящим током в течение всего периода вегетации.
В более поздних исследованиях Хачидзе и Мосашвили (1973) установили в пасоке глюкозу, фруктозу и в виде следов — сахарозу. Из органических кислот в пасоке обнаружены яблочная, винная, лимонная и фумаровая кислоты. Попов и Кръстева (1977) нашли в пасоке аминокислоты, органические кислоты и сахара. Установлено некоторое влияние фосфорного удобрения на органические вещества в пасоке.
Jако (1977), Skene, Antcliff (1972) и ряд других авторов установили в пасоке цитокинины.
Содержание органических веществ в пасоке отмечалось также в более ранних исследованиях.