Сроки закладки соцветий в зимующих почках по длине побега и плодоносность почек. Сроки закладки соцветий в зимующих почках винограда изучали многие авторы.
Наиболее ранние исследования принадлежат Мартелли (Martelli, 1892) и Мюллер-Тургау (Muller-Turgau, 1892). Они показали, что зачатки первичных гроздей можно наблюдать еще в августе и что дифференциация соцветий начинается, когда почки находятся в зеленом состоянии. В Калифорнии (Дэвис) первые признаки дифференциации плодоносных почек в нижних частях побегов отмечены в начале июня. До конца июня соцветия сформировались в почках 15-го глазка (Winkler, Shemsettin, 1937).
Таблица 20
Развитие междоузлий в зачатке побега (в делениях окуляра микрометра)
(1954—1956 гг.)
Дата взятия образца | Длина зачатка побега по зонам | Число междоузлий в среднем на почку | Развитие междоузлий по периодам, | ||
1—4-й узел | 5—7-й узел | 8—10-й узел | |||
15/V...... | 3,1 | 2,6 | _ | 2,8 | 22.4 |
30/V...... | 5.1 | 5,4 | 3,7 | 4,7 | 15,2 |
15/VI...... | 9,5 | 11,4 | 12,4 | 11,1 | 51,2 |
15/VII...... | 10,5 | 12,5 | 13,1 | 12,0 | 7,2 |
15/VIII..... | 11,0 | 12,8 | 13,4 | 12,4 | 3,2 |
15/XII...... | 11,1 | 12,8 | 13,5 | 12,5 | 0,8 |
В Австралии и Южной Америке дифференциация соцветий начинается с середины октября и продолжается до начала декабря (Barnard, 1932; Barnard, Thomas, 1933; Perold, 1927); во Франции начинается примерно с середины июня и продолжается до середины августа (Huglin, 1958), а в Италии начинается и заканчивается примерно на 20—30 дней раньше— 16—28 мая (Breviglieri, 1956).
Исследования 3. Я. Титовой-Молчановой (1951, 1952) в условиях Средней Азии показали, что закладка зачатков соцветий в почках, сортов Пино черный, Саперави, Ркацители. Ак кишмиш и Тайфи розовый начинается почти одновременно (во второй половине мая) и совпадает с периодом цветения винограда.
К аналогичным выводам приходит также А. А. Менагаришвили (1957), который изучал закладку соцветий в глазках сорта Ркацители в условиях Грузинской ССР. Н. П. Науменко и 3. В. Колесник (по сообщению Корнейчука и Плакиды, 1959) также установили, что зачатки первых соцветий изучаемого ими большого набора сортов (Анжевин Оберлен, Шасла доре, Каберне, Гаме черный, Сенсо, Серексия, Рислинг, Алиготе, Ркацители, Пино серый) образуются почти одновременно.
3. Я. Титова-Молчанова (1951, 1952) установила, что у сортов, у которых рост побегов прекращается раньше (Пино черный, Ркацители и Саперави), зачатки первых соцветий закладываются и более сжатые сроки — примерно в течение месяца (заканчивается процесс в конце мая — первой половине июня). У сортов с более длинным вегетационным периодом (Тайфи розовый и Ак кишмиш) закладка зачатков соцветий несколько отстает — наиболее интенсивно этот процесс происходит в конце июня. На основании этого автор приходит к выводу, что периоды наиболее интенсивной закладки
зачатков в почках у разных сортов не совпадают.
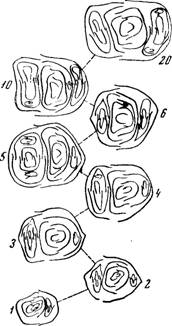
Рис. 20. Поперечные срезы глазков сорта Тайфи розовой (цифрами обозначены порядковые номера глазков по длине побега)
Степень дифференциации и плодоносности почек по длине побега неодинаковы. В средней зоне побега (5—10-й узел) соцветия лучше дифференцированы и почки более плодоносны, чем в нижней. Глазки в средней зоне побега крупные по размеру, в них больше замещающих почек, а в главных почках этих глазков на зародышевом побеге больше узлов и лучше развитых зачатков соцветий (рис. 20).
А. Ф. Тютюник (1952), изучавший развитие зимующих почек винограда и закладку в них соцветий в средней полосе СССР, пришел к выводу, что разница в сроках закладки и дифференциации соцветий в центральной почке десятого глазка у различных сортов довольно значительна и достигает 20—40 дней (табл. 21).
Из приведенных данных также видно, что от закладки бугорка до дифференциации первого
Таблица 21
Сроки закладки и дифференциации соцветий в центральной почке десятого глазка у разных сортов

соцветия проходит около месяца (23—33 дней). Период дифференциации второго соцветия несколько короче (19—30 дней). Закладка бугорка и дифференциация второго соцветия начинается примерно на 10—12 дней позже, чем первого.
Таблица 22
Динамика закладки соцветий в центральных почках глазков в 1952 г.
(в 96)
Узел (зона побега) | 25/У | 9/У1 | 20/У1 | 30/У1 | 11/VIII |
Димят | |||||
1-й | 15 | 35 | 30 | 40 | 52 |
3-й | 18 | 56 | 58 | 74 | 78 |
6-й | — | 36 | 57 | 83 | 95 |
10-й | — | 15 | 55 | 73 | 100 |
15-й | — | — | — | 60 | 94 |
Памид | |||||
1-й | 15 | 40 | 65 | 77 | 83 |
3-й | 17 | 66 | 67 | 76 | 100 |
6-й | _ | 55 | 67 | 87 | 100 |
10-й | — | 19 | — | 75 | 95 |
15-й | — | — | — | 65 | 100 |
Болгар
| |||||
1-й | 0 | 32 | 30 | 31 | 56 |
3-й | 0 | 44 | 45 | 45 | 71 |
6-й | — | 33 | 66 | 67 | 93 |
10-й | — | 17 | 67 | 74 | 95 |
15-й | — | — | — | 61 | 95 |
Мавруд
| |||||
1-й | 0 | 24 | 32 | 40 | 69 |
5-й | 0 | 33 | 46 | 60 | 80 |
6-й | — | 9 | 42 | 65 | 84 |
10-й | —. | — | 48 | 60 | 92 |
15-й | — | — | — | 21 | 100 |
В условиях Болгарии закладка бугорка начинается в последней декаде мая, а соцветий — в конце мая — начале июня (Стоев, Ников, 1956). Разница в сроке наступления закладки соцветий ранних и поздних сортов в начале периода цветения составляет примерно 10—14 дней. Через месяц она почти теряется. Особо характерен в этом отношении сорт Димят. По сроку созревания ягод он принадлежит к почти поздним сортам, однако цветение у него начинается рано, соцветия закладываются тоже рано (табл. 22).
Чтобы установить темпы закладки соцветий по периодам процесса, авторы сопоставили количество соцветий, сформировавшихся в начальные 20 и последние 40 дней (табл. 23).
Таблица 23
Количество соцветий (в %), сформировавшихся в почках в различных зонах побега в период с 9 по 30 июня и с 1 июля по II августа

* К 9 июня соцветий не было.
В начальный период (9—30/VI) за более короткий срок закладывается больше соцветий, чем позже (1/VII—11/VIII) за более длительный срок. К аналогичным выводам приходит также А. А. Менагаришвили (1957), отмечая, что максимальное количество соцветий образуется в период цветения до первой половины июня. В фазу горошения закладывается много соцветий, но все же меньше, чем раньше.
Таким образом, закладку соцветий нельзя рассматривать как процесс, протекающий равномерно в течение вегетационного периода. В 60—65% зимующих почек 15-го узла сортов Димят, Памид и Болгар соцветия заложились всего за период около 10 дней. Наиболее интенсивно процесс идет в средней и выше средней части побега.
Аналогичную закономерность отмечает 3. Я. Молчанова О953), которая установила, что формирование почек происходит тем быстрее, чем выше от основания побега они находятся. В соответствии с этим и дифференциация соцветий в них осуществляется быстрее и лучше, в результате чего эти почки более плодоносны.
Рост и дифференциация соцветий продолжается и после периода интенсивной закладки. Исследования К. Д. Стоева и М. М. Никова (1956) показали, что находящиеся у основания побега глазки вначале имеют более крупные соцветия, а несколько позже значительно усиливается рост соцветий в почках, расположенных выше, поэтому уже к середине августа наиболее развитыми оказываются соцветия в зоне 10— 15-го узла (табл. 24).
Таблица 24 Величина соцветий по длине побега
Узел | Величина соцветий, мк | Узел | Величина соцветий, мк | ||
30/VI | 11/VIII | 30/VI | 11/VIII | ||
| Димят |
|
| Болгар |
|
1-Й . . | 350 | 472 | 1-Й ... . | ._ | 588 |
3-й . . | 504 | 672 | 3-й .... | 336 | 616 |
6-й . . | 574 | 700 | 6-й .... | 392 | 728 |
10-й . . | 392 | 910 | 10-й .... | 386 | 882 |
15-й . . | 224 | 910 | 15-й .... | — | 896 |
| Памид |
|
| Мавруд |
|
1-й . . | 364 | 386 | 1-й ... . | 294 | 546 |
3-й . . | 532 | 784 | 3-й ... . | 560 | 630 |
6-й . . | 518 | 812 | 6-й .... | 392 | 854 |
10-й . . | 350 | 798 | 10-й .... | 350 | 840 |
15-й . . | 280 | 728 | 15-й .... | — | 840 |
Более крупные размеры соцветий в почках средней и выше средней зоны побега установлены многими авторами в различных районах виноградарства и у самых разнообразных сортов. Это можно считать одной из наиболее характерных закономерностей биологии винограда. Наблюдаемые позже, но время вегетации, различия в величине гроздей в разных зонах прошлогоднего побега существуют еще в глазках во время закладки и дифференциации соцветии.

Рис. 21. Коэффициент плодоносности разных сортов винограда 11 августа:
1 — Димят; 2 — Памид; 3 — Болгар; 4 — Мавруд
Вскоре после формирования первых соцветий в почках происходит закладка вторых и последующих. По данным К. Д. Стоева и М. М. Никова (1956) для этого требуются 20—25 дней, причем этот срок длиннее для низко расположенных глазков и короче для расположенных в средней части побега и выше.
О заложении соцветии в замещающих почках сообщает Е. К. Плакида (1953). В ее опытах удаление центральных почек совершенно не сказалось на величине урожая и среднем весе грозди сорта Шасла, но в сильной степени отразилось на урожайности Муската белого.
Способность к формированию нескольких соцветий в почках ярче выражена в глазках, расположенных выше, поэтому коэффициент плодоносности их больше (рис. 21). Лучшая дифференциация соцветий, по-видимому, объясняется лучшим питанием почек благодаря повышенной ассимиляционной активности листьев средних частей основного и бокового побега (Стоев, Занков, Панайотов, Найденова, 1952; Стоев, Добрева, Зейналов, 1965, 1966).
Закладка соцветий происходит также в зимующих почках на пасынках. Как показали исследования К. Д. Стоева и М. М. Никова (1956), закладка соцветий в почках на пасынках начинается в первой декаде июля, причем раньше на пасынках нижних и средних ярусов основного побега (1—4-й и 7—9-й узлы), а позже (в последней декаде июля) и на пасынках выше средней зоны основного побега. По длине пасынка в нижней части его (1—4-й узел) обнаружена акро-петальная последовательность закладки соцветий. На пасынках в зоне 7—9-го узла основного побега плодоносность почек не меняется метамерно, а в зоне 12—15-го узла — увеличивается базипетально.
Авторами установлено также, что на пасынках процент плодоносных почек несколько ниже, чем на основном побеге. Это не совсем согласуется с часто наблюдаемым в производственных условиях более высоким коэффициентом плодоносности пасынков. Несоответствие это можно объяснить тем, что микроскопическим путем не всегда удается точно учесть соцветия в почках пасынков, которые, как правило, меньше, чем в почках основного побега. Можно предполагать также, что заложенные летом бугорки в почках пасынков развиваются дальше и дифференцируются в ранневесенний период или наблюдения закончены были слишком рано (1/IX).
После искусственного проращивания зимующих почек в год их формирования на выросших побегах также образуются сложные почки (глазки), а в них закладываются нормальные соцветия. К. Д. Стоев и М. М. Ников (1956), удаляя все точки роста на побеге после 15 июня, установили возможность закладки соцветий на основном побеге, на развитых на нем пасынках, на основных побегах второй генерации, выросших преждевременно из зимующих почек, и на пасынках второй генерации.
Несколько позже М. Ников (1964), продолжая эту работу, успел получить в течение одного вегетационного периода четыре генерации плодоносных побегов из зимующих почек (последние три генерации за счет преждевременно проросших зимующих почек). При этом в зимующих почках на побегах всех генераций закладывались соцветия.
Таким образом, способность винограда формировать почки на основных побегах и на пасынках и закладывать в них соцветия очень велика. Это имеет большое теоретическое и практическое значение. Факт закладки четырех генераций плодоносных почек объясняется еще тем, что этот процесс в последующих генерациях осуществляется значительно более интенсивными темпами и в более сжатые сроки.
В благоприятных условиях нормальные соцветия закладываются и в почках жирующих побегов, развивающихся из спящих почек, и побегов из замечающих почек глазка.
Вопрос о возможности закладки нормальных плодоносных почек на жирующих побегах сравнительно новый. В последние годы все больше утверждается взгляд, что почки жирующих побегов не менее плодоносны, чем все остальные на побегах, развитые из прошлогодней древесины. Например, А. Ф. Турянский (1950) приводит данные, из которых видно, что нет существенной разницы в плодоносности почек побегов, выросших из главных и замещающих почек основного побега, и побегов, выросших из спящих почек. Н. И. Гузун и В. С. Семин (1963) даже установили наиболее высокую плодоносность почек на побегах, выросших из спящих почек.
Существенный вклад в изучение вопроса о плодоносности почек на жирующих побегах внес Юглен (Huglin, 1955). Он изучал среднее число соцветий в почках жирующих побегов в условиях полного удаления однолетних и двухлетних побегов и при совместном выращивании побегов из спящих почек и нормальных из почек прошлогоднего побега. Наблюдения показали, что нет никакой разницы в плодоносности почек, развивающихся на нормальных и жирующих побегах.
На основании этих результатов автор делает заключение, что почки жирующих побегов имеют такую же плодоносность, как и нормальных побегов с аналогичной силой роста.
Основные закономерности развития соцветий. А. Л. Подражанский (1944) отмечал, что если на побеге одна гроздь, то она, при одинаковых других условиях, всегда весит меньше, чем ниже расположенная на побеге с двумя гроздями А. М Негруль (1948) объясняет это положение существованием критических периодов в развитии зимующих почек. Если условия благоприятствуют закладке лишь одного соцветия в почке, то они чем-то не соответствуют его оптимальному развитию. Значительно позже было установлено, что между числом и размерами зачатков соцветий существует высокая прямая корреляция (Antcliff, Webster, 1955).
По мнению А. М. Негруля (1959), «первый критический период в развитии почки наступает, когда смещенная точка роста, продолжая рост, может образовать или усик, или соцветие. Заложится зачаток соцветия или почка останется бесплодной, зависит в этот момент от внутренних и внешних условий.
В дальнейшем наступает период роста этой почки, который определяет степень ветвления и сложность соцветия, т. е. его величину и количество бутонов. С этим периодом тесно связаны и последующие, в которые закладываются зачатки второго и третьего соцветий. Все эти периоды приходятся на разное время для разных почек и наступают по мере их формирования по длине побега, обычно в июне — сентябре».
В литературе по биологии винограда (Баранов, 1946; Мержаниан, 1967, и др.) общепризнанным является положение, что соцветие и усик имеют генетически одинаковое происхождение (симподиальная теория). Симподиальный рост побегов винограда характерен тем, что у 3—5-го узла побега на конусе нарастания в пазухе листа возникает бугорок, который начинает расти очень интенсивно и, опережая в росте другие секторы конуса нарастания, превращается в ведущий ростовой побег. Верхушечная точка нарастания главного побега, отставая в росте, отклоняется в сторону и, регенерируя, превращается в соцветие или усик.
Как показали наблюдения, в зонах 3—8-го узла симподиальное отклонение превращается, как правило, в соцветие, а в более высоко расположенных зонах — в усик. Можно считать вполне доказанной закономерность, что выше последнего соцветия всегда закладываются усики, а выше первого усика никогда не бывает соцветий и гроздей. Правда, в литературе отмечены случаи, когда на одном побеге соцветия имеются значительно выше 5—8-го узла. Например, М. М. Ников (1962) наблюдал у сортов Димят и Алиготе по 5 гроздей на одном побеге, а у сорта Япладжа белая — 13 гроздей, находящихся между 3-м и 21-м узлом. А. М. Негруль (1959) отмечал у сорта Тельти куйруги наличие 7 гроздей. М. Г. Цейтлин (1952) указывает, что основная масса соцветий располагается от первого до девятого узла зеленого побега. Наиболее часто грозди развиваются на 3—6-м узлах, реже на 1—2-м и на 7—10-м.
Имея в виду, что в зимующей почке обычно развиваются максимально 12—14 зачаточных узлов, надо полагать, что случаи, когда на побеге обнаруживаются соцветия и грозди слишком высоко (выше допустимой длины зачаточного побега в зимующей почке), являются результатом внепочечной закладки соцветий. О внепочечной закладке соцветий сообщает Г. Ф. Турянский (1950), который наблюдал нормальное развитие соцветия на 26-м и 28-м узлах.
Поэтому, рассматривая проблему о закладке соцветий в зимующей почке, надо отметить, что кардинальным вопросом является механизм превращения первого симподиального отклонения в соцветие или усик. Пока наши познания об этом механизме, как и об условиях, являющихся решающими в превращении соцветия в усик или обратно, очень бедны.
А. М. Негруль (1959) считает, что образованию соцветия из первого симподиального бугорка способствует наличие благоприятных условий. При отсутствии таких условий симподиальное отклонение превращается в усик, и тогда почки, а следовательно и выросшие из них побеги, остаются бесплодными. (Понятие благоприятные условия касается главным образом притока питательных веществ, обеспечивающих нормальный рост зачаточного побега в зимующей почке и закладку соцветий.)
А. М. Негруль и Л. Н. Гордеева (1968) стремились выяснить сроки прохождения первого критического периода, а также расшифровать условия, благоприятствующие формированию плодоносных почек в зимующем глазке винограда. Для этой цели они применяли кольцевание побегов и полное удаление листьев в разные сроки развития почек и на различной высоте побега что должно было, по предположению авторов, нарушить нормальный ход формирования главной почки в зимующем глазке и уменьшить процент ее плодоносности. Чтобы установить, на каких этапах жизнедеятельности эмбрионального побега недостаток питания является наиболее неблагоприятным фактором, с 25 июня по 1 сентября проводили анатомо-морфологические исследования состояния меристемы конуса нарастания центральных почек зимующих глазков в момент формирования плодовых бугорков. По состоянию почек средней зоны побега (пятая и шестая почки от основания побега) авторы определяли степень влияния неблагоприятных условий питания на закладку плодовых бугорков и дальнейшую их дифференциацию в соцветия.
1 Варианты опыта: I — кольцевание побега и удаление листьев у 5—6-го междоузлия 26 мая; II — те же операции проведены 5 июня при длине побега 7—8 междоузлий; III—те же операции проведены 16 июня при длине побега 9—10 междоузлий; IV — те же операции проведены 26 июня при длине побега 11—13 междоузлий; V — контроль, свободный рост побегов с прищипкой пасынков над 2—3-м листом.
Результаты наблюдений показали, что условия, созданные на первых этапах органогенеза зачаточного побега (варианты I, II, III), привели к тому, что бугорки соцветий или не заложились совсем (вариант I) или заложились, но образовались усики или усиковые соцветия (варианты II и III). Из этого следует, что условия, созданные кольцеванием и полным удалением листьев на ранних этапах роста побегов, являются неблагоприятными. Они ухудшили питание, в результате чего был нарушен нормальный ход развития зимующих почек — в них не заложились соцветия и они сильно отстали в росте.
Кольцевание и полное удаление листьев, проведенные позднее при развитии 11—13-го междоузлий на побеге, привели к некоторому отставанию в росте зимующих почек (их. размеры немного меньше по сравнению с почками контрольного варианта), но все же в них заложились плодовые бугорки и произошла их дальнейшая дифференциация в нормальные соцветия. Таким образом, неблагоприятные условия, созданные на более поздних этапах, немного задержали рост, но все же не помешали качественному переходу к закладке соцветий в зимующих почках.
Вполне нормально проходили рост и развитие почек в контрольном варианте, где не проводили никаких мероприятий, приводящих к ухудшению питания зачаточных побегов. В конце вегетации у них обнаружено по два соцветия, а эмбриональный побег достиг наибольшей величины.
На основании проведенных исследований авторы вновь говорят о наличии критического периода в развитии зимующих почек, который наступает после отчленения меристемой конуса нарастания почки определенного числа вегетативных органов. Для зачатка основного побега (центральной почки) критическое состояние совпадает с образованием 3—4-го листовых зачатков.
Очень важно утверждение авторов о том, что состояние критического момента в формировании почек — явление динамическое, так как оно непрерывно сдвигается от основания побега к его верхушке. Для почки 2—3-го узла, например, критический момент наступает при образовании 6—7 междоузлий побега, для 4—6-го узла — при 9—11, для 7— 10-го узла — при образовании 12—14 междоузлий.
Для прохождения критических периодов, по мнению А. М. Негруля и Л. Н. Гордеевой, необходимым условием является не только приток ассимилятов, но также наличие ряда элементов минерального питания.
Исследования Н. И. Гузуна и В. С. Семина (1963) также показали, что имеется положительная зависимость между дифференциацией почек и соцветий и транслокацией Р32. По данным Рзндольфа (Randolph, 1944), после подкормки фосфором значительно возросло число соцветий в почках сорта Кармен. Аналогичное явление обнаружено также под влиянием удобрения цинком.
Поскольку оптимальные условия питания не наступают одновременно по всей длине побега, закладка плодоносных бугорков и дифференциация соцветий в почках не проходит равномерно. В конечном счете качество почек (плодоносность, а по всей вероятности и жизненность) по длине побега неодинаково.
Многие авторы, изучавшие проблему плодоносности почек винограда, отмечают, что в почках средних зон побега больше крупных соцветий (Турянский, 1950; Молчанова, 1953; Негруль, Гордеева, 1968; Стоев, Ников, 1956; Царев, 1952; Мельник, 1953; Тавадзе, 1958; Корнейчук, Плакида, 1959; Khalil, 1961; Ников, 1962; Коваль, Никифорова, 1962; Мелконян, 1964; Гузун, Литвак, 1964, и др.). Не разбирая подробно данные различных авторов, отметим, что повышение плодоносности зимующих почек от основания к средним и выше расположенным узлам побега, наблюдающееся хотя и не со строгой математической последовательностью,— закономерность биологии развития винограда. Это, как считают все авторы, связано с условиями питания и среды, сочетание которых является предпосылкой значительных метамерных изменений плодоносности зимующих почек винограда.
По нашему мнению, способность винограда закладывать более развитые соцветия в вышерасположенных зонах побега генетически обусловлена. К этому выводу мы пришли, изучая плодоносность почек, сформированных на побегах, выросших из глазков в год их формирования при принудительном проращивании (Стоев, Ников, 1956). Несмотря на то что побеги выросли в конце июня — начале июля, а формирование зимующих почек и закладка соцветий в них произошли в основном в августе и отчасти в сентябре, т. е. при наличии лучшего сочетания условий, чем весной, все же способность винограда формировать более плодоносные почки в средних зонах побега сохранилась (табл.25).
Таблица 25
Узел | Димят | Болгар | Юни блан | |||
% плодоносных почек | величина соцветий, мк | % плодоносных почек | величина соцветий, мк | % плодоносных почек | величина соцветий, мк | |
1-й..... | 80 | 602 | 60 | 532 | 64 | 448 |
3-й..... | 100 | 700 | 64 | 672 | 85 | 560 |
6-й..... | 100 | 812 | 72 | 644 | 85 | 644 |
10-й..... | 100 | 798 | 80 | 630 | 90 | 602 |
Все это свидетельствует о том, что свойство винограда формировать больше лучше развитых соцветий в почках средних зон побега по сравнению с расположенными ниже является биологической закономерностью, выработанной в процессе эволюции. Поэтому объяснение, что недостаточное развитие соцветий в низкорасположенных почках побега — результат неблагоприятных условий внешней среды во время их формирования, как нам кажется, не раскрывает полностью, а может быть и действительную причину этого явления.
Идея о существовании критических периодов в развитии и дифференциации соцветий зимующих почек винограда по длине побега правильная. Однако вопрос об оптимальных для прохождения критических моментов условиях, а тем более с применением соответствующих методов регулирования питания растения для обеспечения наиболее благоприятных условий закладки плодоносных побегов в зимующих почках нельзя считать решенным. Нередко при наличии казалось бы благоприятных условий закладка соцветий происходит не совсем нормально, в результате чего на следующий год урожай бывает не очень высоким. Конечно, вопрос значительно осложняется тем, что дифференциация зачаточных бугорков при наличии благоприятных условий может продолжаться и в ранневесенний период (Колесник, 1953, 1959). По наблюдениям И. Н. Кондо, В. В. Крылова и А. И. Литвака (1964), с завершением фазы глубокого покоя рост эмбриональных побегов может возобновляться и длина их может несколько увеличиваться (табл. 26). Происходит это в основном не вследствие деления клеток верхушечной меристемы почек и образования новых узлов и междоузлий, а за счет вставочного (интеркалярного) роста, т. е. разрастания эмбриональных клеток ранее сформированных зачаточных междоузлий и узлов.
Таким образом, некоторое увеличение длины центральной почки глазков, наблюдаемое с помощью микроскопа, после завершения фазы глубокого покоя происходит главным образом в результате последовательного растяжения эмбриональных клеток в акропетальном направлении — сначала нижних междоузлий, а затем средних и верхних междоузлий и узлов почки.