Наиболее опасные грибные заболевания винограда милдью (возбудитель Plasmopara viticold), оидиум (возбудитель Uncinula necator) и виноградная тля филлоксера (Phylloxera vastatrix) были завезены в Европу из Северной Америки во второй половине прошлого столетия. Быстро распространившись на виноградниках европейских стран, они стали причинять большой вред, так как культивировавшиеся в Европе сорта винограда, относящиеся к виду Vitis vinifera, оказались совершенно неустойчивыми к этим болезням и вредителю. Естественно, начались поиски устойчивых форм. Было установлено, что дикие американские виды винограда устойчивы к филлоксере, милдью и оидиуму.
А. М. Негруль (1959) объясняет устойчивость диких американских видов и неустойчивость европейских сортов винограда их филогенетическим положением. Родиной филлоксеры, милдью и оидиума является юго-восточная часть Северной Америки, где они приспособились к питанию на винограде. В результате длительной эволюции при постоянном контакте паразиты и виноград взаимно приспосабливались и изменялись, чему способствовали благоприятные экологические условия.
ФИЛЛОКСЕРОУСТОЙЧИВОСТЬ
Корни и листья всех видов винограда рода Vitis, которые культивируются на виноградниках, поражаются филлоксерой. Согласно общеизвестной двадцатибалльной шкале Раваза и Виала, филлоксероустойчивость отдельных видов оценивается следующим образом: американские виды V. rupestris — 19,5 балла, V. riparia — 19 баллов, V. solonis, V. berlandieri — 15 баллов, V. labrusca — 5 баллов, европейские сорта V. vinifera — 0—1 балла.
Европейские сорта винограда обладают высоким качеством плодов, но они неустойчивы к филлоксере. Некоторые из них, например Ркацители, Мцване, Серексия, Греческий розовый и др., могут дольше противостоять вредоносному действию филлоксеры, чем Шасла, Саперави, Мускат белый. Алиготе и др. У европейских сортов винограда филлоксера сильно повреждает корни, которые в местах поражения отмирают и загнивают. Вследствие этого кусты находятся в угнетенном состоянии или гибнут. На листьях европейских сортов под влиянием всех биотипов филлоксеры образуются некрозы (некротические пятна, «блюдца», стерильные галлы). В районах сплошного заражения филлоксерой европейские сорта винограда прививают на филлоксероустойчивых подвоях.
Для получения филлоксероустойчивых подвоев путем гибридизации используют американские виды V. riparia, V. rupestris, V. solonis, V. berlandieri. Они устойчивы к филлоксере, но плоды их непригодны для употребления в пищу. Корни упомянутых видов поражаются филлоксерой, но на них происходит полное заживление поврежденных тканей. Поэтому корни продолжают нормально функционировать и кусты хорошо растут. На листьях американских видов и их гибридов филлоксера не всегда вызывает образование галлов.
В 20—30-х годах настоящего столетия в Западной Европе проходила большая дискуссия. Она началась с работ Бернера (Вoгпег, 1914, 1923, 1924, 1925), который утверждал, что в Европе существуют две расы филлоксеры, северная (Phylloxera vastatrix) и южная (Phylloxera vitifolii). Северная раса, распространенная в Германии, не поражала в Лотарингии листья и корни некоторых сортов американских видов, в то время как южная поражала их. Данные Бернера встретили возражения ряда ученых, которые касались главным образом вопроса константности этих рас (Grassi, 1924; Topi, 1929; Schneider-Orelli, Stellwaag, 1924).
Дискуссия вызвала ряд новых исследований, осветивших вопросы биологии и особенно экологии филлоксеры и ее способности поражать листья американских видов. На большом числе примеров было показано, что в зависимости от экологии и условий питания вирулентность филлоксеры может изменяться, что поставило под сомнение существование бернеровских рас как константных форм. На западноевропейских виноградниках наблюдалась потеря вирулентности филлоксеры, длительное время питавшейся на европейских сортах винограда. Такая филлоксера теряла способность поражать листья и корни американских видов. Маршалл (1923) предполагает, что филлоксера Ph. vastatrix, которая не поражала некоторые сорта американских видов, является потомством особей, размножавшихся в течение многих лет на европейских сортах без участия половых генераций. Я. И. Принц (1937) наблюдал такое же явление в Кахетии и Азербайджане.
На юге Одесской области на листьях сорта Рупестрис дю Ло, относящегося к V. rupestris, местный биотип филлоксеры не образует галлов, но филлоксера, привезенная из района города Могилева-Подольского, вызывает галлообразование на всех зараженных ею растениях. Н. Н. Троицкий (1932) сообщает, что на листьях Рупестрис дю Ло галлы хорошо развиваются при заражении местными биотипами в Туапсе, в Кахетии и на Сакарской опытной станции. Из сказанного можно заключить, что у филлоксеры существуют расы, отличающиеся по вирулентности, и селекционеры при выведении филлоксероустойчивых сортов винограда должны считаться с этим фактом.
Первая работа о причине филлоксероустойчивости винограда опубликована Мильярде (Millardet, 1898). В ней было показано, что в корнях американских устойчивых видов пораженные филлоксерой ткани отделяются слоем опробковевших клеток. Отслаивается всего лишь несколько слоев клеток луба и после их отделения корень остается совершенно здоровым. У неустойчивых европейских сортов винограда отслаивающий слой пробковых клеток не образуется и ткани корня отмирают на большую глубину, после чего начинается их загнивание. Теория Мильярде нашла всеобщее признание, но она не объясняла причины образования пробкового слоя и не затрагивала вопроса о поражаемости листьев филлоксерой.
В 1919 г. Мольц (Molz, 1919) опубликовал работу, в которой утверждал, что филлоксероустойчивые лозы винограда обладают большой кислотностью клеточного сока. При проверке оказалось, что такой коррелятивной зависимости не существует.
Ряд авторов связывают филлоксероустойчивость с особым анатомическим строением корней (Абесадзч и др., 1930; Кискин, 1957; Berner, 1942). Однако исследования такого рода не вскрыли причину образования на листьях галлов или некрозов, и, кроме того, анатомические различия между корнями устойчивых и неустойчивых сортов винограда были замечены на многолетних корнях, а на мочковатых корешках первичного строения различий не найдено. Несмотря на это, мочковатые корешки при поражении филлоксерой устойчивых американских видов остаются живыми, а у сортов неустойчивых погибают (Зотов, 1955).
Я. И. Принц (1951, 1965) и А. В. Благовещенский (1945, 1950) выдвинули фенольную теорию филлоксероустойчивости винограда. Они утверждают, что филлоксера не может питаться на сортах, содержащих много фенолов, так как у нее происходит инактивация протеолитического фермента слюны.
А. В. Благовещенский (1950) предположил, что этот протеолитический фермент относится к группе катепсинов. Хиноны, образующиеся при окислении флавонов и фенолов, действуют как окислители на активный катепсии, переводят его в недеятельное состояние, вследствие чего филлоксера лишается возможности питаться продуктами глубокого распада белков. Однако филлоксера питается и нормально размножается на корнях устойчивых американских видов в течение всего вегетационного периода. Этого не могло бы происходить, если ферменты ее слюны инактивировались.
Исследования В. В. Зотова и Т. И. Соколовской (1959) показали, что фенолов больше всего у самого неустойчивого к филлоксере сорта Саперави и что корни различного возраста содержат разное количество фенолов, а в мочковатых корнях они практически отсутствуют. На основании сказанного можно сделать вывод, что первоначальное количество фенолов, содержащихся в корнях, не может быть причиной филлоксероустойчивости.
Ф. Штельвааг-Киттлер (Slellwaag-Kittler, 1955) сообщил, что, по его наблюдениям, способность филлоксеры поражать мочковатые корешки зависит от длины корневого чехлика. По его утверждению, филлоксера делает укол только над корневым чехликом, длина которого изменяется в зависимости от сорта н почвенных условий. Корни сортов с коротким чехликом восприимчивы, а с длинным устойчивы к корневой форме филлоксеры. В. В. Зотов наблюдал, что филлоксера делает укол через корневой чехлик, и его длина, по-видимому, не играет большой роли, так как на разнообразных почвах Украины, Молдавии и Грузии на мочковатых корнях европейских сортов винограда и американских видов в изобилии образуются корневые галлы.
Андерс (Anders, 1960) считает, что некоторые свободные аминокислоты слюны филлоксеры могут стимулировать образование галлов на листьях винограда. В связи с этим Генке (Непке, 1963) связывает филлоксероустойчивость со способностью тканей листьев винограда дезаминировать аминокислоты слюны филлоксеры, вследствие чего галлы не могут развиваться. О. Генке считает, что интенсивность дезаминирования аминокислот зависит от количества хинонов, образующихся в результате окисления фенолов полифенолоксидазой, которые затем дезаминируют аминокислоты слюны филлоксеры. Генке брал для анализов кружочки листьев диаметром 1 см, следовательно, листья были большие. Филлоксера же может вызвать образование галлов, если она делает укол в верхушечный, еще не полностью раскрывшийся листочек. Таким образом, О. Генке исследовал ткани листа в таком возрасте, в котором они не могут быть использованы для изучения специфической ответной реакции на поражение филлоксерой.
Кроме того, этот автор не изучал ответной реакции на корнях. Между тем хорошо известно, что на мочковатых корешках образуются галлы как на устойчивых, так и на неустойчивых к филлоксере сортах винограда.
Изучение явления филлоксероустойчивости изолированно на корнях и на листьях может привести к искаженным представлениям о филлоксероустойчивости куста винограда в целом. В действительности так и получалось. Ряд ученых (Berner, 1933; Троицкий, 1929; Принц, 1951, 1965, и др.), наблюдая образование нормальных галлов на листьях американских видов винограда и некрозов на европейских сортах, противопоставляли неустойчивость к филлоксере листьев устойчивости корней. Нет сомнения, что правильное представление о филлоксероустойчивости может быть получено лишь в том случае, если это явление изучают на всем кусте, а не на отдельных его органах.
Морфологические изменения корней и листьев, вызванные филлоксерой, выражаются в следующем. На мочковатых корнях филлоксера делает укол вблизи меристемы в то место, где заканчивается дифференциация клеток и начинается их растяжение. После этого на мочке образуется утолщение, т. е. корневой галл с углублением в том месте, где присосалась филлоксера.
У устойчивого сорта после образования корневого галла мочка продолжает нормально расти (рис. 86, 2) и остается нормальной толщины. При дальнейшем росте на ней может образоваться еще несколько корневых галлов подобно бусам, нанизанным на шнурок. Ветвление мочковатого корешка продолжается нормально. Корневые галлы покрываются опробковевшей покровной тканью, остаются живыми в течение всего вегетационного периода и перезимовывают. Весной клетки, образующие опухоль, отслаиваются, а корешок остается живым. Отмирать могут лишь те корневые галлы, которые образовались на мочках, останавливающихся в росте и отмирающих естественным путем, независимо от того, есть на них корневой галл или нет.
У неустойчивого сорта рост мочки, на которой образовался корневой галл, приостанавливается (рис. 86, 3). Кончик корневого галла в 2—2,5 раза толще нормальной мочки. Ветвление мочковатых корешков становится ненормальным, уродливым. Корневые галлы неустойчивых сортов скоро погибают. У сорта Шасла массовое их отмирание происходит на 45—60-й день. Отдельные экземпляры гибнут раньше или позже. К осени большинство мочковатых корешков, на которых образовались корневые галлы, гибнет, и кусты страдают от недостатка влаги даже в том случае, если в почве ее достаточно. В осенне-зимний период все поздно образовавшиеся корневые галлы отмирают.

Рис. 86. Образование галлов на корнях винограда при поражении филлоксерой:
1 — здоровый мочковатый корешок винограда; 2—галлы на корнях устойчивого сорта Рипария X Рупестрис 101-14; 3 — галлы на корнях неустойчивого сорта Шасла
На одревесневших корнях образуются опухоли (туберозитеты), которые по размеру и форме соответствуют колонии филлоксеры. По мере увеличения колонии увеличивается и размер опухоли. На устойчивых сортах опухоли мало заметны, поздней осенью или в конце весны следующего года они легко очищаются, и под ними обнаруживается молодая покровная ткань. На неустойчивых сортах опухоли вздуты, в начале заболевания имеют белесый цвет и под ними либо совсем не образуется покровной ткани, либо она образуется частично. К осени или весне следующего года на этих местах открываются незаживающие или частично заживающие раны.
Нормальные листовые галлы образуются на листьях устойчивых сортов американских видов. Их вызывают отродившиеся личинки первого возраста, которые поднимаются на верхушку побега и делают укол на молодом не полностью раскрывшемся листочке с верхней стороны листовой пластинки. Вокруг места сосания филлоксеры ткани начинают разрастаться и через 10—12 дней образуется нормальный галл (рис. 87, г). На 12—14-й день присосавшаяся внутри галла филлоксера начинает откладывать яйца. Одна особь может отложить до 500 яиц, после чего отмирает (естественно). Ткань галла остается живой до листопада.
На листьях неустойчивых к филлоксере европейских сортов в местах уколов образуются некрозы: некротические пятна, «блюдца» и стерильные галлы с отмершей в центре тканью (рис. 87, а, б, в). Формирование галла прекращается потому, что вместе с отмиранием ткани гибнет филлоксера и действие ее слюны прекращается. На 7—10-й день после начала отмирания ткани получается некротическое пятно, через 10—12 дней «блюдце». Если отмирание ткани начинается позже чем на 12—14-й день после того, как филлоксера достигает возраста взрослой самки, образуется стерильный галл. Стерильные галлы отличаются от нормальных тем, что в них нет филлоксеры, они меньше и имеют заостренное побуревшее «дно».

Рис. 87. Повреждения филлоксерой листьев неустойчивых европейских сортов (а, б, в) и устойчивых американских видов (г) винограда: а — некротическое пятно, б — «блюдце»; в — стерильный галл; г — нормальный галл
В редких случаях на листьях отдельных кустов европейских сортов могут образоваться галлы, по своим признакам приближающиеся к нормальным. Филлоксера живет в них дольше (до 20—25 дней) и успевает отложить небольшое количество яиц, но затем гибнет, так как ткань вокруг места ее сосания отмирает, возникает некроз и получается стерильный галл позднего образования. Поскольку такие явления наблюдаются на некоторых кустах и отдельных побегах, мы связываем их с процессом клонообразования. Есть указания, что на листьях европейских сортов, растущих в оранжереях, также развиваются галлы, но они стерильные, различных типов и среди них есть приближающиеся к нормальным. Кроме того, на листьях кустов, растущих в оранжереях, одновременно с галлами образуются и блюдца.
Филлоксера — сосущее насекомое, относится к подотряду тлей. Патологические изменения в органах винограда, вызванные филлоксерой, происходят под воздействием ее слюны. Колющий аппарат филлоксеры состоит из четырех щетинок, которые она складывает попарно, образуя две капиллярные трубочки. По одной из них она вводит слюну, по другой — всасывает растворы питательных веществ, подготовленные внекишечным пищеварением и готовые к усвоению. Таким образом, действие слюны на клетки пораженных тканей направлено на превращение их содержимого в питательный субстрат. Быстрое разрастание галловой ткани связано с тем, что излюбленным местом укола филлоксеры являются ткани вблизи меристемы быстро растущих органов, богатых РНК, к которым обеспечен постоянный сильный приток пластических веществ: кончики мочковатых корешков, первый верхушечный листочек, одревесневшие корни вблизи камбия.
Некоторые авторы считают (Принц, 1965), что у филлоксеры внекишечное пищеварение настолько совершенно, что подготовленный питательный субстрат полностью без остатка усваивается и поэтому кишечник у филлоксеры закрытый.
Биохимическим свойствам слюны филлоксеры посвящен ряд работ, на основании которых можно составить представление о ее действии на пораженные ткани винограда.
Бернер считает, что в слюне филлоксеры содержится цецидогенное вещество, стимулирующее образование галлов. Если вместо галла образуется некроз, то это объясняется тем, что в тканях имеются антитела (антицецидогенные вещества). Цвайгелт (Zweigelt, 1941) отрицает наличие стимуляторов в слюне филлоксеры и считает, что она своим действием активизирует комплекс веществ, содержащихся в клетках пораженных тканей, побуждая их к разрастанию. Образование галлов Цвайгелт рассматривает как активную защитную реакцию.
Клофт (Klaft, 1951) и Гопп (Норр, 1955) нашли в слюне филлоксеры стимуляторы роста, наличием которых и объясняют образование галлов на корнях и листьях. Природы этих веществ они не установили, но подтверждают их наличие опытами с корнями кресс-салата, которые под действием слюны филлоксеры давали вздутия. Кроме того, они приводят в доказательство всем известный факт, что рост галлов прекращается, если филлоксера удалена, и, следовательно, нет притока слюны к пораженной ткани.
Андерс (Anders, 1960) установил, что в слюне филлоксеры ауксины отсутствуют, но нашел в ней пять свободных аминокислот: триптофан, гистидин, лизин и, по-видимому, глютаминовую кислоту и валин. Ф. Андерс не без основания считает, что свободные аминокислоты в слюне филлоксеры играют роль возбудителя разрастания ткани и образования галлов. Особенно интересно присутствие триптофана, который, как известно, является предшественником ауксинов.
Нужно отметить, что Л. А. Собецкий и М. А. Державина (1963) нашли в слюне филлоксеры до 16 свободных аминокислот. Это расхождение с результатами исследований Ф. Андерса следует объяснить тем, что Л. А. Собецкий и М. А. Державина брали для исследования взрослую, давно питавшуюся филлоксеру, и поэтому в экстракт попадал не только секрет слюнных желез, но и содержимое пищеварительного тракта.
Б. А. Рубин, Е. В. Арциховская и Т. В. Денисова (1964), исследуя слюну еще не питавшихся личинок филлоксеры, не нашли в ней ИУК, но обнаружили четыре аминокислоты — гиетидин, лизин, глютаминовую «ислоту и триптофан. Так как триптофан и гистидин представляют собой соединения индольного характера, авторы испытали действие их слабых растворов на корни сеянцев винограда и получили экспериментальное опухолеобразование. На этом основании они пришли к выводу, что гистидин и триптофан принимают участие в активации ростовых процессов в тканях винограда, пораженных филлоксерой.
Установлено, что в слюне филлоксеры содержатся весьма активные протеолитические ферменты и амилаза (Зотов, 1955), расщепляющие соответственно белки и крахмал. Путем гистохимических исследований было также показано, что у места непосредственного сосания филлоксеры в клетках разрушается РНК (Зотов, 1964). Это говорит о том, что в слюне филлоксеры содержится активная рибонуклеаза.
Имеющиеся в литературе сведения о биохимических свойствах слюны филлоксеры не столь обширны, как бы этого хотелось, но то, что уже известно, с достаточным основанием позволяет сделать вывод о том, что слюна филлоксеры оказывает двоякое действие на ткани винограда. С одной стороны, при помощи гидролитических ферментов она деполимеризует сложные соединения, превращая содержимое клеток в питательный субстрат, а с другой — оказывает стимулирующее действие на ростовые процессы. Это подтверждается тем, что опухолевые клетки отличаются по направленности биохимических процессов (Зотов, 1964; Арциховская, Денисова, 1966). У места сосания филлоксеры возникает зона, в которой преобладают процессы гидролиза содержимого клеток (зона гидролиза), а в прилегающих клетках превалируют процессы синтеза (зона синтеза). Эти зоны легко обнаруживаются при окрашивании препаратов йодом для выявления крахмала, места образования которого служат как бы индикатором, показывающим зоны, в которых преобладают процессы полимеризации (Зотов, 1964; Зотов, Светлякова, Соколовская и др., 1966). Развитие болезни, как на листьях, так и на корнях, зависит от устойчивости сорта.
В случае устойчивости в корневых галлах, вокруг зоны гидролиза образуется зона синтеза (рис. 88). Поздней осенью или ранней весной вблизи эндодермы и по краям галловой ткани возникает феллоген, отслаивающий опухолевые клетки, а корешок остается живым. У неустойчивого сорта зона синтеза образуется очень слабо или совсем отсутствует и в конце второго месяца корневые галлы гибнут вместе с мочковатыми корешками, на которых они образовались. Кусты лишаются всасывающих корней.
На одревесневших корнях филлоксероустойчивого сорта под зоной гидролиза вскоре возникает зона синтеза. На их разделе образуется слой клеток, обогащенных РНК; они начинают делиться и получается феллоген, отслаивающий небольшие участки пораженной ткани. Происходит как бы самоизлечение. На неустойчивых к филлоксере сортах зона гидролиза распространяется по всей толще луба и в древесину. Феллоген не образуется, и в местах повреждения филлоксерой на корнях возникают большие открытые раны.
В тканях листьев, пораженных филлоксерой, наблюдается то же, что и в корнях. На листьях устойчивых сортов она вызывает развитие нормальных галлов. В них вокруг зоны гидролиза возникает зона синтеза, т. е. развивается синтетическое направление обмена. Галловая ткань при этом остается живой до листопада. В листьях же неустойчивого сорта зона синтеза вокруг зоны гидролиза не образуется. На 7- 12-й день пораженная ткань отмирает и образуется некроз.
Такого рода различия в ответной реакции устойчивого и неустойчивого сортов винограда обусловливаются интенсивностью и направленностью биохимических процессов, наступающих в тканях растения в ответ на попадание в них слюны филлоксеры.

Рис. 88. Анатомические изменения в корнях и листьях устойчивого (А) (Рипария X Рупестрис 101—14) и неустойчивого (Б) (Шасла) сортов, пораженных филлоксерой:
1 - корневые галлы; 2 — одревесневшие корни (у устойчивого сорта видно отслаиваний тканей у неустойчивого — отмирание); 3 — листья (у устойчивого сорта образуется нормальный галл, у неустойчивого происходит отмирание тканей и образуются некрозы различных типов); а — зоны гидролиза (место сосания филлоксеры); б — зона синтеза; в — зона растяжения; г — слой клеток, богатых РНК; 1 — феллоген, ОМ — остаточная меристема
Об этих изменениях можно судить по накоплению сложных соединений в галловой ткани, которых у устойчивых сортов синтезируется гораздо больше, чем у неустойчивых (Зотов, 1964) (табл. 119).
Таблица 119
Содержание полимерных соединений в здоровых мочках и корневых галлах (в мг на 1 г сырого вещества)
| Сорт | Ткань | Азот щелочнорастворимого белка | Крахмал |
Шасла (неустойчивый) | Здоровые мочки . . . | 1,04 | 0,6— 0,7 |
Корневые галлы . . . | 1,87 | 20,3— 69,5 | |
Рипария X Рупестрис | Здоровые мочки. . . | 1,03 | 0,4— 0,6 |
101—14 (устойчивый) | Корневые галлы . . . | 2,63 | 88,7—109,6 |
Об интенсивности синтеза в корневых галлах можно судить по накоплению сухих веществ. В здоровых мочках содержится около 100 мг сухих веществ на 1 г сырого вещества. В корневых галлах неустойчивого сорта Шасла к концу четвертой декады от начала поражения их накапливается до 135—185 мг сухих веществ. Однако в дальнейшем накопление их прекращается, и корневые галлы вскоре гибнут. Нарастание количества сухих веществ в корневых галлах устойчивого сорта Рипария X Рупестрис 101—14 продолжается в течение всего вегетационного периода. К концу вегетации в них содержится 240—280 мг сухих веществ, и галлы остаются живыми (Зотов, 1964).
Белковый обмен, являющийся центральным процессом жизнедеятельности клеток, безусловно, лежит в основе всех изменений, наступающих в пораженных филлоксерой тканях винограда. В настоящее время установлено, что ДНК, РНК и белок составляют единую взаимосвязанную систему. Изменение белкового обмена в тканях винограда, пораженных филлоксерой, связано с нарушением обмена нуклеиновых кислот. Гистохимические исследования показали, что филлоксера разрушает РНК в зоне гидролиза.
Как показали исследования Р. И. Светляковой и др. (1966), в 45—50-дневных корневых галлах неустойчивого сорта Шасла перед началом их отмирания содержание РНК остается почти без изменения, а количество ДНК удваивается; отношение РНК к ДНК уменьшается почти в 2 раза (табл. 120). У филлоксероустойчивого сорта Рипария X Рупестрис 101—14 в тот же срок содержание РНК в корневых галлах возрастает в 3,5, а ДНК более чем в 1,5 раза. Отношение РНК к ДНК увеличивается в 2 раза.
В преднекротических пятнах листьев сорта Шасла, по данным гистохимических исследований, содержание РНК уменьшается, так как в полости клеток, особенно в столбчатой паренхиме, пиронинофилия резко ослабевает. Количество ДНК не изменяется потому, что ядра сохраняют прежние размеры, окрашиваются метиловым зеленым и дают фельген-положительную реакцию. Надо полагать, что отношение РНК к ДНК становится меньше, чем в здоровых клетках. В тот же срок в листовых галлах сорта Рипария X Рупестрис 101—14 содержание РНК возрастает почти в 4, а ДНК более чем в 1,5 раза. Отношение РНК к ДНК увеличивается в 2,2 раза.
Таблица 120
Содержание РНК и ДНК в здоровых и пораженных филлоксерой листьях (в мг на 100 см2 листовой пластинки) и корнях винограда (в мг на I г сырой ткани)

* Преднекротические пятна имеют незначительные размеры, поэтому анализировали их только гистохимическн. При этом отмечено уменьшение РНК, а количество ДНК не изменяется.
Таким образом, в пораженных тканях винограда на единицу ДНК у филлоксероустойчивого сорта синтезируется в 2 раза больше РНК, а у неустойчивого в 2 раза меньше по сравнению со здоровой тканью. Это находится в прямой связи с особенностями биосинтеза белка в больной ткани.
Аминокислотный состав белков в тканях винограда, пораженных филлоксерой, резко изменяется (табл. 121, 122).
Как видно из табл. 121 и 122, в белках корней найдено 18, а в белках листьев — 16 аминокислот.
После поражения филлоксерой в корневых и листовых галлах филлоксероустойчивого сорта Рипария X Рупестрис 101 —14 содержание аминокислот увеличивается, но неравномерно. По цистеину и оксипролину наблюдается даже небольшое снижение, а содержание триптофана почти не изменяется.
Таблица 121
Отношение содержания аминокислот белков корневых галлов к белкам здоровых почек
(по Соколовской, 1966)
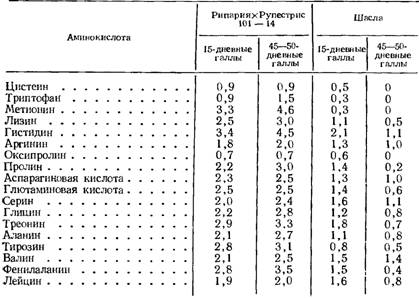
Таблица 122
Отношение содержания аминокислот белков листовых галлов и преднекротических пятен к белкам здоровых листьев
(по Соколовской, 1966)
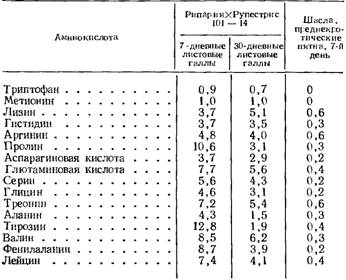
Количество остальных аминокислот в белках корневых галлов (табл. 121) увеличивается в 2—4,5 раза. В этом отношении особенно выделяются метионин и гистидин. В листовых галлах различия выражены еще больше, особенно на седьмой день поражения (табл. 122), причем в этот период содержание аминокислот в белках возрастает больше, чем в 30-дневных листовых галлах. Это говорит о том, что у филлоксероустойчивого сорта включение аминокислот в белки листовых галлов происходит особенно интенсивно в начале заболевания. Приведенные данные показывают, что в случае филлоксероустойчивости в тканях винограда, пораженных филлоксерой, усиливается биосинтез белка. Изменение соотношения аминокислот в этих белках свидетельствует о том, что их состав в количественном отношении изменяется. По всей вероятности, происходит также биосинтез специфических белков, свойственных только пораженным филлоксерой тканям.
У неустойчивого к филлоксере сорта Шасла в пораженных тканях листьев и корней наблюдается снижение интенсивности включения аминокислот в белки (табл. 121, 122). Уже в 15-дневных корневых галлах значительно снижается содержание цистеина, триптофана, метионина, оксипролина и тирозина. Почти не изменяется количество лизина, глицина и аланина, а содержание остальных аминокислот повышается в 1,5—2 раза. Но уже в 45—50-дневных корневых галлах перед началом отмирания включение аминокислот в белки начинает прекращаться: цистеин, триптофан, метионин и оксипролин в этот период заболевания не были обнаружены в белках и только количество гистидина, аргинина, аспарагиновой кислоты, серина и валина остается близко к норме (табл. 121). В преднекротических пятнах на седьмой день содержание аминокислот в белках составляет всего лишь 0,2—0,6 нормы, а триптофан и метионин совсем не обнаружены (табл. 122). Прекращение включения некоторых аминокислот в белки больных тканей неустойчивого к филлоксере сорта показывает, что на определенном этапе заболевания биосинтез целого ряда белков не происходит, в том числе и каталитических.
Примером белка-фермента, биосинтез которого прекращается, может быть цитохромаксидаза. Е. В. Арциховская и Т. В. Денисова (1966) наблюдали, что в галлах неустойчивого к филлоксере сорта Шасла активность цитохромоксидазы резко падает, а перед началом отмирания пораженной ткани этот фермент в галлах совсем не удается обнаружить. В то же время в галловой ткани филлоксероустойчивого сорта Рипария X Рупестрис 101 —14 количество клеток, дающих положительную реакцию на цитохромоксидазу, увеличивается в 2,5—3 раза.
Не менее интересные данные получены по активности полифеноксидазы и пероксидазы (Зотов, 1964; Арциховская, Денисова, 1966). У устойчивого сорта винограда в корневых галлах, вызванных филлоксерой, активность указанных ферментов остается на более высоком уровне, чем в здоровой ткани на протяжении всего вегетационного периода, в то время как у неустойчивого она резко падает ниже нормы, и корневые галлы отмирают. По данным тех же авторов, система полифенолы — полифенолоксидаза играет большую роль в филлоксероустойчивости винограда. Активность полифенолоксндазы в пораженных корнях возрастает, коррелируя с увеличением содержания фенольных соединений, которых у устойчивого сорта Рипария X Рупестрис 101—14 накапливается гораздо больше по сравнению с неустойчивым. Например, у сорта Шасла уже в 3—4-й декаде от начала заболевания накопление полифенолов прекращается. Роль системы полифенолы — полифенолоксидаза в защитной реакции может быть связана, в частности, с образованием барьеров из продуктов необратимого окисления фенолов и, по-видимому, с частичной инактивацией ферментов слюны филлоксеры.
Цитогистохимические исследования показали, что описанные выше изменения метаболизма клеток, пораженных филлоксерой, приводят к изменению их метаболитов (Зотов, 1964). Так, в клетках зоны синтеза корневых галлов филлоксероустойчивого сорта Рипария X Рупестрис 101—14 ядра увеличиваются в 8 раз, а ядрышки — в 7 раз по сравнению со здоровыми, а у сорта Шасла соответственно — в 4 и в 2 раза. Еще большие различия наблюдаются в листьях. Если в зоне синтеза листовых галлов Рипария X Рупестрис 101—14 ядра увеличиваются в 20 раз, а ядрышки в 30 раз, то у сорта Шасла в клетках, прилегающих к зоне гидролиза, они остаются почти такого же размера, как в здоровых листьях.
Увеличение размера ядер и ядрышек сопровождается образованием гранул. У устойчивого к филлоксере сорта они возникают в ядре (рис. 89). По мере созревания гранулы перемещаются к его периферии и отделяются от него; во время отделения они окрашиваются янусом зеленым. Следовательно, в них протекают окислительные процессы. В отделившихся гранулах начинает накапливаться крахмал и они превращаются в крахмальные зерна. Этот процесс продолжается в течение всего вегетационного периода. В клетках зоны синтеза корневых галлов устойчивого сорта всегда много отделяющихся от ядер гранул, а вся полость клетки заполняется крахмальными зернами.
У неустойчивого к филлоксере сорта гранулы образуются путем отпочковывания от ядерной оболочки (рис. 90). В них во время отделения начинают формироваться кристы, а под световым микроскопом можно наблюдать, что они окрашиваются янусом зеленым. Отпочковывание гранул от ядерной оболочки скоро прекращается.

Рис. 89. Электронно-микроскопическая фотография клеточного ядра из зоны синтеза корневого галла устойчивого сорта Рипария X Рупестрис 101—14. В ядре видны образующиеся гранулы (показаны стрелками).
ЯК — ядрышко
Сделанные наблюдения говорят о том, что от клеточных ядер отделяются дыхательные гранулы, причем в корневых галлах устойчивого сорта этот процесс протекает интенсивно на протяжении всего лета, а у неустойчивого сорта очень слабо и лишь в начале заболевания.
Во фракции ядер и отделяющихся от них гранул было определено окислительное форсфорилирование (Зотов, 1964). Эти опыты показали, что при поражении филлоксерой в корневых галлах устойчивого сорта винограда активация дыхания сопровождается усилением фосфорилирования, в то время как у неустойчивого сорта наблюдается разобщение этих процессов и окислительное фосфорилирование оказывается полностью подавленным на пятой декаде от начала заболевания (табл. 123). Такие различия следует объяснить тем, что в пораженных тканях устойчивого сорта продолжается биосинтез белков и в ядрах образуются гранулы в течение всего вегетационного периода. У неустойчивого сорта вследствие
глубокого нарушения белкового обмена образование гранул прекращается.
В начале заболевания дыхание корневых галлов сорта Шасла усиливается почти в 2 раза (Зотов, Светлякова, Соколовская и др., 1966), а затем резко снижается и в конце второго месяца галлы начинают отмирать. Интенсивность дыхания корневых галлов Рипария X Рупестрис 101 —14 возрастает почти в 1,5 раза и затем на протяжении всего лета и начала осени остается на более высоком уровне по сравнению со здоровыми мочками (табл. 124). Корневые галлы при этом остаются живыми.

Рис. 90. Электронно-микроскопическая фотография клеточного ядра из зоны синтеза корневого галла неустойчивого сорта Шасла. Образования гранул в ядре не происходит. От оболочки ядра отделяются гранулы (показаны стрелками), в них видны крнсты. ЯК — ядрышко
Таблица 123
Окислительное фосфорилирование во фракции клеточных ядер и отделяющихся от них гранул

Таблица 124
Интенсивность дыхания корневых галлов и здоровых мочек (в мкатомах 02, поглощенного за 1 ч 1 г сырой ткани)

Таким образом, в клетках пораженной ткани филлоксероустойчивых форм винограда, в противоположность неустойчивым, происходит приспособление дыхательного аппарата к новым условиям, складывающимся под воздействием филлоксеры, в результате чего дыхательные функции и их энергетические параметры не только сохраняются, но в значительной степени усиливаются. Это подтверждает теоретическую концепцию, развиваемую Б. А. Рубиным (1960), о том, что исход заболевания растений, всегда связанный с изменением интенсивности окислительных процессов в пораженных тканях, зависит от степени использования энергии дыхания. У устойчивых форм освобождающаяся при этом энергия аккумулируется в форме макроэргических фосфатных связей, которые используются для различных форм защитных реакций, прежде всего для усиления синтетического направления обмена и подавления гидролизующего действия ферментов.
Накопленный экспериментальный материал позволяет понять взаимосвязь между процессами, протекающими в тканях листьев и корней, пораженных филлоксерой, и различие между устойчивыми и неустойчивыми сортами винограда.
У устойчивого сорта усиление синтеза РНК и белков, повышение общего уровня энергетического обмена обеспечивают развитие синтетического направления обмена и жизнь клеток в тканях, пораженных филлоксерой. Это благоприятствует возникновению феллогена, который отслаивает на корнях небольшие участки отмершей ткани. Питавшаяся в них филлоксера при этом погибает. В результате в тканях, находящихся под феллогеном, обмен продолжается, корни остаются живыми и заболевание оканчивается самоизлечением. Эти же процессы на листьях устойчивого сорта в местах сосания филлоксеры приводят к образованию нормальных галлов, ткани которых остаются живыми до осеннего листопада.
У сорта винограда, неустойчивого к филлоксере, утраченная РНК не восстанавливается, биосинтез белков слабеет, нарушается энергетический обмен и усиливается гидролизующее действие слюны филлоксеры. Вполне понятно, что в таких условиях не может возникнуть образовательная ткань — феллоген, поэтому на корнях образуются большие открытые раны. Эти же процессы на листьях неустойчивого сорта в местах сосания филлоксеры приводят к возникновению некрозов.
Изложенное выше свидетельствует о том, что ответная реакция на поражение филлоксерой, возникающая в тканях корней и листьев, характеризуется тождественными биохимическими процессами, характер которых зависит от степени устойчивости сорта, а морфологические признаки пораженных филлоксерой листьев отражают процессы, происходящие в пораженных тканях. На этой основе разработан метод определения филлоксероустойчивости по типу поражения листьев (Зотов, 1967).
Сущность этого метода заключается в том, что после искусственного заражения листьев филлоксерой отбирают те формы винограда, на листьях которых образуются только нормальные галлы, что соответствует полному заживлению корней в местах поражения филлоксерой. Выше было сказано, что отдельные расы филлоксеры отличаются по своей вирулентности. Поэтому для заражения листьев нужно подбирать наиболее агрессивные ее биотипы.
Существуют также методы определения филлоксероустойчивости по типу поражения корней. Они разработаны Т. И. Соколовской (1961) и П. X. Кискиным (1962). Суть методов заключается в том, что с зараженных филлоксерой кустов винограда путем раскопок отбирают пробы корней, на которых определяют интенсивность образования феллогена в местах поражения филлоксерой. У устойчивых форм на поврежденных филлоксерой корнях образуется сплошной феллоген, полностью отделяющий пораженные участки ткани.
Применяют еще так называемый прямой метод определения филлоксероустойчивости, заключающийся в том, что кусты заражают филлоксерой с тем, чтобы она уничтожила неустойчивые формы. Хотя этот метод распространен, от него следует отказаться. Дело в том, что неустойчивые к филлоксере европейские сорта винограда в некоторых случаях могут довольно долго выдерживать вредоносное действие филлоксеры, что зависит от следующих причин.
На развитие филлоксеры влияет механический состав почвы и ее физические свойства. Чем больше кварцевых частиц, тем малочисленнее филлоксера на корнях винограда. В почвах, содержащих глинистых частиц не более 4—5%, филлоксера не живет. Она так же плохо развивается на заиленных наносных почвах. Благоприятны для развития филлоксеры структурные черноземы (Казас, Гаркавенко, Пойченко, 1960; Принц, Иванов, 1948).
Агротехнические условия влияют на сопротивляемость филлосексере кустов неустойчивых европейских сортов винограда (Мордвилко, 1925; Зотов, 1955; Казас, Гаркавенко, Пойченко, 1960; Михайлов, 1960). Все, что снижает синтетическую функцию в корнях, ускоряет гибель кустов, зараженных филлоксерой. Мощные формировки, допускающие накопление на кустах многолетней древесины, недогрузка кустов урожаем, поливы, регулярное внесение удобрений, подкормки и пр.— все это повышает сопротивляемость кустов. Напротив, малые формировки без многолетней древесины, перегрузка кустов урожаем, частые дефициты влаги в летние месяцы, истощенная почва — снижают сопротивляемость филлоксере неустойчивых сортов винограда. Время заражения также играет большую роль. В случае заражения двухлетних кустов, вредоносное действие филлоксеры может быть обнаружено на 7—8-й год. Если они заразятся в 3—5-летнем возрасте, то состояние явного угнетения может наступить значительно позже, на 10—15-й год. В этом случае нетрудно допустить ошибку, сделав преждевременный вывод.
Более продолжительное выживание кустов некоторых европейских сортов винограда, зараженных филлоксерой, П. Н. Недов (1965) объясняет тем, что их корни после повреждения этим вредителем гниют с различной интенсивностью. У некоторых сортов, по его утверждению, гниение настолько медленное, что кусты долгое время не гибнут. П. Н. Недов указывает, что в различных почвенных условиях корни одних и тех же европейских сортов винограда после повреждения филлоксерой подвергаются гниению то в большей, то в меньшей степени, что обусловливается двумя факторами: наличием инфекционного запаса в почве и условиями, которые в ней складываются для развития гнилостных процессов.
Таким образом, перечисленные факторы в определенных сочетаниях могут затушевывать истинную филлоксероустойчивость сортов винограда и селекционных форм.
Иногда даже 15-летнего срока недостаточно, чтобы сделать правильный вывод. Ошибочно отобранные формы в другом месте, при иных условиях, могут погибнуть от филлоксеры после первого же плодоношения, т. е. на 4—5-й год после посадки.
Не трудно видеть, что основные факторы, мешающие определению филлоксероустойчивости винограда, связаны с почвенными условиями. Поэтому наиболее точно и просто можно определить устойчивость к филлоксере по типу поражения листьев. Большую точность дают так же упомянутые методы Т. И. Соколовской (1961) и П. X. Кискина (1962). Однако они отличаются сложностью и громоздкостью в связи с необходимостью раскопок корневой системы каждого куста и последующего анатомо-гистохимического анализа отобранных образцов корней.