В. В. Зотов, Р. Ш. Гадиев
УСТОЙЧИВОСТЬ ВИНОГРАДА К ВРЕДИТЕЛЯМ И БОЛЕЗНЯМ
Наиболее опасные грибные заболевания винограда мильдью (возбудитель Plasmopara viticola Berl. et de Toni), оидиум (возбудитель Uncinula necator (Scw.)Burr) и виноградная тля филлоксера (Phylloxera vastatrix) были завезены в Европу из Северной Америки во второй половине прошлого столетия, быстро распространившись на виноградниках европейских стран, они стали причинять большой вред, так как культивировавшиеся в Европе сорта винограда, относящиеся к виду Vitis vinifera, оказались совершенно неустойчивыми к этим болезням и вредителю. Естественно, начались поиски устойчивых форм. Было установлено, что дикие американские виды винограда устойчивы к филлоксере, мильдью и оидиуму.
Негруль (1959) объясняет устойчивость диких американских видов и неустойчивость европейских сортов винограда их филогенетическим положением. Родиной филлоксеры, мильдью и оидиума является юго-восточная часть Северной Америки, где они приспособились к питанию на винограде. В результате длительной эволюции при постоянном контакте паразиты и виноград взаимно приспособились и изменились, чему способствовали благоприятные экологические условия. На этой основе происходил естественный отбор.
ФИЛЛОКСЕРОУСТОЙЧИВОСТЬ
Корни и листья всех видов рода Vitis, которые культивируются на виноградниках, поражаются филлоксерой. Только два вида — Vitis rotundifolia и Vitis cinerea Arnold иммунны к этому вредителю, но они не используются в хозяйственных целях из-за ряда отрицательных свойств. Причина их иммунитета заключается, по-видимому, в том, что филлоксера не может ими питаться. Здесь природу филлоксероустойчивости этих видов рассматривать не будем.
Американские виды V. riparia, V. rupcstris, V. solonis, V. Bcrlandieri, V. Labrusca и др. устойчивы к филлоксере, но им присуще низкое качество ягод. Европейские сорта винограда V. vinifera обладают высоким качеством ягод, но они неустойчивы к филлоксере. Некоторые из них, например, Ркацители, Мцване, Серексия, Греческий розовый и др., могут дольше противостоять вредоносному действию филлоксеры, чем сорта Шасла, Саперави, Мускат белый, Алиготе и др.
При первичном заражении филлоксера поселяется на всех корнях, но излюбленным местом ее питания являются мочковатые корешки.

Рис. 1. Филлоксерные повреждения листьев
а — нормальный галл; б — стерильный галл; в — блюдце; г — некротическое пятно.
Увел. 6 раз
Личинка филлоксеры стремится сделать укол в ткани, где происходит деление клеток в зону растяжения мочковатых корешков первичного строения и в первый верхушечный листочек.
На мочках устойчивых сортов образуются мелкие опухоли — корневые галлы, которые, если корешок принимает вторичное строение, остаются живыми все лето и перезимовывают. На следующий год в мае — июне опухолевая ткань корневых галлов отмирает, к этому сроку раневая перидерма хорошо сформировывается и полностью защищает центральный цилиндр корня. Пока опухолевая ткань живая, корневые галлы плотно заселены питающейся на них филлоксерой; несмотря на это, мочковатые корешки не гибнут, хорошо растут и ветвятся. Лишь небольшое число вредителя переселяется на толстые одревесневшие корни. На них появляются мелкие опухоли (туберозитеты), ткань которых остается живой все лето.
Под него образуется раневая перидерма. Если она успевает сформироваться к осени, то отмирание опухоли и ее отслаивание происходит в октябре или в ноябре. Иногда этот процесс затягивается до мая — июня следующего года. На листьях филлоксероустойчивых сортов развиваются нормальные галлы (рис. 1), в которых самки филлоксеры питаются до двух месяцев. Галловая ткань на листьях, как и на корнях, остается живой в течение всего лета, но существование листовых галлов прерывается осенним листопадом. Такая взаимосвязь типов поражения листьев и корней на сортах винограда, устойчивых к филлоксере, показывает, что при вегетативном размножении филлоксероустойчивые корни вырастают из черенков, взятых с лоз, на листьях которых образуются нормальные галлы.
На мочковатых корнях неустойчивых к филлоксере европейских сортов винограда разрастаются крупные опухоли — корневые галлы, превосходящие толщину мочки в пять — шесть раз. При небольшом урожае, хорошем уходе за кустами корневые галлы остаются живыми до двух месяцев, но некоторые начинают отмирать уже через месяц. Вследствие столь быстрой гибели корневых галлов происходит отмирание значительной части мочковатых корешков, и филлоксера переселяется на одревесневшие толстые корни. На них она вызывает образование глубоко проникающих в ткани крупных опухолей, которые через один — два месяца отмирают. Под отмершей опухолевой тканью обнажается древесина, не защищенная раневой перидермой. Отмершая ткань загнивает, филлоксера переселяется на новые здоровые участки корней, быстро расселяясь по ним до тех пор, пока не погубит всю корневую систему куста. Таким образом, опухоли на корнях неустойчивых сортов винограда гибнут быстро, поэтому при раскопках давно зараженных филлоксерой кустов обращают на себя внимание не сами опухоли, а их последствия, то есть гниющие участки корней. Если ограничиться этими поверхностными наблюдениями и не изучать физиолого-биохимические изменения, наступающие в пораженной филлоксерой ткани, то может сложиться ложное представление, что корни гибнут вследствие гниения, а не под действием филлоксеры.
На листьях неустойчивых европейских сортов винограда ткань, пораженная филлоксерой, быстро отмирает на всю толщину мозофилла. Уже через 10—20 дней после укола тли на листьях можно наблюдать стерильные галлы, блюдца и некрозы, прекратившие свое развитие на ранних стадиях образования с отмершей тканью в центре (рис. 1). Следовательно, ткани листьев и корней неустойчивых сортов быстро отмирают под действием слюны филлоксеры. Поэтому при вегетативном размножении неустойчивые сорта винограда из черенков, взятых с лоз, на листьях которых не образуются нормальные филлоксерные галлы, имеют и неустойчивые к филлоксере корни.
На листьях американских видов и их гибридов филлоксера не всегда вызывает образование галлов. В 20—30-х годах настоящего столетия в Западной Европе происходила большая дискуссия. Она началась с работ Бернера, который утверждал, что в Европе существуют две расы филлоксеры: северная (Phylloxera vastatrix) и южная (Phylloxera vitifolii), которые обладают различной вирулентностью. Например, северная раса, распространенная в Германии, вызывает на листьях Рупестрис дю Ло образование галлов, а на Рипариа х Рупестрис 3309 под ее влиянием возникают некрозы. Южная филлоксера, напротив, индуцирует на листьях Рипариа х Рупестрис 3309 галлы, а на Рупестрис дю Ло — некрозы. Данные Бернера встретили возражения ряда ученых, которые касались, главным образом, вопроса константности этих рас (Grassi, 1924; Topi, 1929; Schneider-Orelli, Leuzinger, 1924; Ste11waag, 1924). На большом числе примеров было показано, что в зависимости от экологии и условий питания вирулентность филлоксеры может изменяться, что поставило под сомнение существование бернеровских рас как константных форм. На западноевропейских виноградниках наблюдалась потеря вирулентности филлоксеры, длительное время питавшейся на европейских сортах винограда. Такая филлоксера теряла способность поражать листья и корни американских видов. Маrсhall (1923) предположил, что филлоксера Ph. vastatrix, которая не поражала некоторые сорта американских видов, является потомством особей, размножавшихся в течение многих лет на европейских сортах без участия половых генераций. Принц (1937) наблюдал такое же явление в Кахетии и Азербайджане.
На юге Одесской области на листьях сорта Рупестрис дю Ло, относящегося к V. rupestris, местный биотип филлоксеры не образует галлов, но филлоксера, привезенная из района Могилева-Подольского, вызывает галлообразование на всех зараженных ею растениях. Троицкий (1932) сообщает, что на листьях Рупестрис дю Ло галлы хорошо развиваются при заражении местными биотипами на Кавказе.
Первая работа о причине филлоксероустойчивости винограда опубликована Millardet (1898). В ней было показано, что в корнях американских видов пораженные филлоксерой ткани отделяются слоем опробковевших клеток. Отслаивается всего лишь несколько слоев луба, и после их отделения корень остается совершенно здоровым. У неустойчивых европейских сортов винограда отслаивающий слой пробковых клеток не образуется и ткани корня отмирают на большую глубину, после чего начинается их загнивание. Теория Мильярде нашла всеобщее признание, но она не объясняла причины образования пробкового слоя и не затрагивала вопроса о поражаемости листьев филлоксерой.
Ряд авторов связывает филлоксероустойчивость с особым анатомическим строением корней (Абесадзе с сотр., 1930; Кискин, 1957; Вorner, 1942), что, конечно, имеет некоторое основание. Однако исследования рода не вскрыли причину образования раневой перидермы на корнях и галлов на листьях. Кроме того, анатомические различия между корнями устойчивых и неустойчивых сортов винограда были замечены на многолетних корнях, а на мочковатых корешках первичного строения различий не найдено. Несмотря на это, мочковатые корешки при поражении филлоксерой устойчивых американских видов остаются живыми, а у сортов неустойчивых погибают (Зотов, 1955).
Филлоксера — сосущее насекомое, относится к подотряду тлей. Она может питаться только в живой ткани. Патологические изменения в органах винограда, вызванные ею, выражаются в образовании опухолей (галлов), в которых ее личинка при помощи вводимой ею слюны создает специфические условия, благоприятные для питания филлоксеры. Слюнные железы филлоксеры простираются от передней части тела до среднегруди, что указывает на их большой объем, обеспечивающий обильное выделение слюны (Rilling, 1967). Колющий аппарат филлоксеры состоит из четырех щетинок, которые она складывает попарно, образуя две капиллярные трубочки. По одной из них она вводит слюну, по другой всасывает растворы питательных веществ, подготовленные внекишечным пищеварением к усвоению. Некоторые считают (Принц, 1965), что у филлоксеры внекишечное пищеварение настолько совершенно, что подготовленный питательный субстрат полностью без остатка усваивается, и поэтому кишечник у филлоксеры закрытый.
Биохимическим свойствам слюны филлоксеры посвящено много работ. Вorner (1933) считает, что в слюне филлоксеры содержится цецидогенное вещество, стимулирующее образование галлов. Если вместо галла образуется некроз, то это объясняется тем, что в таких тканях имеются антитела (антицецидогенные вещества). Zwеigell (1941) отрицает наличие стимуляторов в слюне филлоксеры и считает, что она своим действием активизирует комплекс веществ, содержащихся в клетках пораженных тканей, побуждая их к разрастанию. Образование галлов на листьях Цвайгелт рассматривает как активную защитную реакцию. Kloft (1951) и Норр (1966) нашли в слюне филлоксеры стимуляторы роста, наличием которых и объясняют образование галлов на корнях и листьях. Природу этих веществ они не установили, но подтверждают их наличие опытами с корнями кресс-салата, которые под действием слюны филлоксеры давали вздутия. Кроме того, они приводят в доказательство всем известный факт, что рост галлов прекращается, если филлоксера удалена, и, следовательно, нет притока слюны к пораженной ткани.
Anders (1960) установил, что в слюне филлоксеры ауксины отсутствуют, но нашел в ней пять свободных аминокислот: триптофан, гистидин, лизин и, по-видимому, глютаминовую кислоту и валин. Ф. Андарс считает, что свободные аминокислоты играют роль возбудителя разрастания ткани и образования галлов. Особенно интересно присутствие триптофана,который, как известно, является предшественником ауксинов. В связи с этим Henke (1963) связывает филлоксероустойчивость со способностью тканей листьев винограда дезаминировать аминокислоты слюны филлоксеры, вследствие чего галлы не могут развиваться. О. Генке считает, что интенсивность дезаминирования аминокислот зависит от количества хинонов, образующихся в результате окисления фенолов полифенолоксидазой, которые затем дезаминируют аминокислоты слюны филлоксеры.
Принц (1951, 1965) и Благовещенский (1950) выдвинули фенольную теорию филлоксероустойчивости винограда. Они утверждают, что филлоксера не может питаться на сортах, содержащих много фенолов, так как у нее происходит инактивация протеолитического фермента слюны. Благовещенский (1950) предположил, что этот протеолитический фермент относится к группе катепсинов. Хиноны, образующиеся при окислении флавонов и фенолов, действуют как окислители на активный катепсин, переводят его в недеятельное состояние, вследствие чего филлоксера лишается возможности питаться продуктами глубокого распада белков. Однако филлоксера питается и нормально размножается на корнях устойчивых американских видов в течение всего вегетационного периода. Этого не могло бы происходить, если ферменты слюны инактивировались.
Исследования Зотова и Соколовской (1959) показали, что фенолов больше всего у самого неустойчивого к филлоксере сорта Саперави и что корни различного возраста содержат разное количество фенолов, а в мочковатых корнях они практически отсутствуют. На основании сказанного можно сделать вывод, что первоначальное количество фенолов, содержащихся в корнях, не может быть причиной филлоксероустойчивости.
Собецкий и Державина (1963) нашли в слюне филлоксеры 16 свободных аминокислот.
Рубин с сотр. (1964), исследуя слюну еще не питавшихся личинок филлоксеры, не нашли в ней ИУК, но обнаружили четыре аминокислоты — гистидин, лизин, глютаминовую кислоту и триптофан. Так как триптофан и гистидин представляют собой соединения индольного характера, авторы испытали действие их слабых растворов на корни сеянцев винограда и получили экспериментальное опухолеобразование. Сторожук (1970) также нашла в слюне филлоксеры триптофан и считает, что эта аминокислота играет роль в галлообразовании, так как приток ее в ткань может повышать содержание β-индолилуксусной кислоты.
Зотов и Гадиев (1975) нашли в ткани галлов, индуцированных филлоксерой, вещества, которых нет в здоровых. Два из них (№1 и 2) обнаружены в слюне личинок филлоксеры. По своим свойствам эти вещества из галловых тканей листьев и корней и слюны личинок филлоксеры идентичны. На хроматограммах они занимают одинаковое положение, в ультрафиолетовом свете флуоресцируют светло-зеленым цветом. Вещество № 1 имеет максимум 266 пт, № 2 — 272—274 пт. Водные растворы этих веществ в физиологических концентрациях стимулировали корнеобразование у черенков фасоли более чем в два раза, у черенков винограда — в три и пять раз. Они повышали всхожесть семян пшеницы и усиливали рост корней на 120—175%. У Botrytis cinerea стимулировалось прорастание спор и развитие мицелия на 150—170%. Эти же авторы нашли в экстрактах из галловой ткани β-индолилуксусную кислоту. Ее не было в вытяжках из слюны личинок филлоксеры, а также из здоровых листьев, закончивших рост. По-видимому, триптофан, поступающий в ткань со слюной, способствует повышению содержания β-индолилуксусной кислоты в галлах.
Таким образом, в слюне личинок филлоксеры содержится комплекс веществ, стимулирующих рост и образование опухолей.
Исследованиями ряда авторов в слюне филлоксеры обнаружены весьма активные протеолитические ферменты, РНК, ДНК и амилаза, расщепляющие соответственно белки, нуклеиновые кислоты и крахмал (Зотов, 1955; Принц, 1951; Anders, 1960; Рубин с сотр., 1964; Сторожук, 1970). Своим исследованиями Сторожук (1970) установила активность ферментов слюны еще не питавшихся личинок филлоксеры (табл. 1).
В первые два дня активность ферментов высокая, на третий день резко падает и личинки гибнут, если они не начнут питаться.
Собецкий и Державина (1966) пытались обнаружить протеазы непосредственно в слюнных железах, для чего при помощи очень тонкого метода выделяли их из филлоксеры. В гомогенате выделенных желез они не обнаружили протеолитической активности. Однако известно, что протеолитические ферменты вырабатываются в виде неактивных белков, которые после выделения превращаются в активные протеазы. Такое приспособление предотвращает самопереваривание тканей, вырабатывающих эти ферменты (Диксон, Уэб, 1961).
Обнаружение в секретах филлоксеры ростовых веществ и протеолитических ферментов раскрывает суть их двоякого действия: стимуляции роста (опухолеобразование), с одной стороны, и лизиса — с другой.
Активность ферментов слюны личинок филлоксеры (один час инкубации)
Дни после рождения | РНК (в относительных единицах) | ДНК (в относительных единицах) | Протеазы (нарастание не белкового азота), mg | Амилаза (нарастание восстанавливающих сахаров), mg |
1-й | 2,25 | 0,15 | 0,56 | 14,7 |
2-й | 1,45 | 0,14 | 0,48 | 18,7 |
3-й | 0,30 | 0,11 | 0,23 | 3,6 |
Итак, в слюне филлоксеры содержатся физиологически активные вещества и гидролитические ферменты (протеазы, РНК, ДНК и амилаза), что вызывает у виноградного растения развитие опухолевых тканей-галлов. При этом ткани корней как устойчивых, так и неустойчивых сортов реагируют на внедрение слюны филлоксеры образованием опухолей, развитие которых, в случае устойчивости приводит на корнях к формированию защитной раневой перидермы, а у неустойчивых сортов — к отмиранию опухолевой ткани с образованием глубоких открытых ран. Причину этого различия нужно искать в особенностях биохимических процессов, развивающихся в галловой ткани.
Schaefer (1974) изучал особенности биохимических процессов в листовых галлах филлоксероустойчивых американских подвойных сортов винограда. Показано, что в начале питания филлоксера оказывает активное влияние на метаболизм биологически важных соединений — белков, нуклеиновых кислот, ферментов в индуцированных ею галловых тканях. После того как происходит естественное отмирание самки филлоксеры и прекращается ее действие, метаболическая активность в галловых и прилегающих тканях приближается к нормальным тканям. Эти очень интересные исследования Н. Schaefer имеют большое значение в понимании влияния филлоксеры на ткани виноградного растения и хорошо согласуются с общеизвестным фактом, что развитие галла прекращается, если удалить филлоксеру.
Раневая перидерма может образоваться только в тканях, клетки которых сохраняют в условиях патогенеза высокую метаболическую активность. Б. А. Рубин с сотрудниками установили, что устойчивым формам растений свойственна высокая прочность сопрягающих систем протопласта, которые в условиях дезорганизующего действия патогенов продолжают функционировать, что лежит в основе защитных реакций, связанных, прежде всего, с повышением общего уровня энергетического обмена и усиления биосинтеза в пораженной ткани (Рубин с сотр., 1975).
Электронно-микроскопические исследования показали, что на первоначальном этапе опухолеобразования, вызванного филлоксерой, происходит активная фрагментация каналов эндоплазматического ретикулума, изменяется конфигурация клеточных ядер, увеличивается количество рибосом на мембранах эндоплазматического ретикулума, в аминопластах образуются крахмальные зерна, которые быстро растут, отодвигая органоиды клеток к периферии. Установлено, что образование опухолей на корнях филлоксероустойчивых сортов винограда происходит лишь в верхних слоях коры. У неустойчивых сортов опухоли охватывают кору, мягкий луб и частично древесину (Чеботарь, 1974).
На примере филлоксероустойчивого сорта Рипариа х Рупестрис 101—14 и неустойчивого сорта Шасла белая были показаны различия в ультраструктуре протопластов клеток в процессе развития опухолевой (галловой) ткани. В опухолевых клетках листьев и корней филлоксероустойчивого сорта сохраняются ультраструктуры протопласта и происходит новообразование клеточных органоидов. Хроматин ядер сохраняет нормальное строение. Ядра увеличиваются в размере, дают рогообразные выросты, на концах которых оболочка разрывается, и между цитоплазмой и ядром устанавливается прямой контакт. В этих местах образуется большое количество митохондрий и пластид. Ядрышки увеличиваются в размере, на периферии у них хорошо выражена нуклеолонемная часть с гранулами, что свидетельствует об усиленном процессе образования рибосом. Эргастоплазма заполняет все промежутки, свободные от крахмальных зерен. Следовательно, протопласт опухолевых клеток на листьях и корнях филлоксероустойчивых сортов винограда сохраняется живым все лето и служит местом питания филлоксеры (Зотов, Клигерман, 1970; Клигерман, Зотов, 1973). В это время под опухолью на корнях формируется раневая перидерма. Поздней осенью или весной следующего года она полностью отслаивает отмирающие опухоли и надежно защищает корень (Зотов, Штеренберг, 1964; Зотов с сотр., 1966). Хотя филлоксера нормально питается и размножается на корнях и листьях устойчивых сортов, кусты хорошо растут. Таким образом, филлоксероустойчивость — это результат взаимного приспособления, при котором филлоксера и виноградное растение уживаются.
На листьях и корнях неустойчивых к филлоксере сортов винограда в опухолевых тканях происходит деструкция ультраструктур. На корнях через 20 дней после укола филлоксеры хроматин клеточных ядер становится волокнистым, плохо контрастируется. В ядрах появляются осмиофильные тяжи — признак разрушения биомембран. Ядро постепенно теряет структуру. Митохондрии набухают, полость клеток заполняется осмиофильными глобулами. Клетки сильно увеличиваются в размере и заполняются большими вакуолями. Все это приводит к отмиранию пораженной ткани на листьях через 8—10 дней, а на корнях — через 1—2 месяца. Ткань отмирает на большую глубину, так как на корнях опухоли захватывают всю кору и значительную часть древесины, а раневая перидерма не образуется (Зотов, Штеренберг, 1964; Чеботарь, 1974).
По данным гистохимических исследований в клетках галловой ткани, в которых накапливаются пластические вещества, увеличивается размер ядер» и ядрышек; у устойчивого сорта Рипариа х Рупестрис 101—14 в 7—8 раз, у неустойчивого сорта Шасла — в 2—3 раза. Вокруг ядер устойчивого сорта наблюдается много мелких органоидов, которые прижизненно окрашиваются янусом зеленым, что свойственно митохондриям (Зотов, 1964; Зотов, Клигерман, 1970). Таким образом, у неустойчивых сортов винограда в тканях листьев и корней под воздействием слюны филлоксеры происходит деструкция клеточных структур, в то время как у филлоксероустойчивых сортов структурная организация протопласта обеспечивает большую прочность клеточных органоидов, которые сохраняют ультраструктуру в условиях неблагоприятного воздействия филлоксеры.
Таблица 2
Процент выделившегося белка (по азоту) из 30-дневных корневых галлов винограда в тpuc-цитратном буфере pH 8,6 без детергента и с детергентом тритоном Х-100

Как известно, клеточные структуры состоят, в основном, из белков, липидов и нуклеиновых кислот, причем молекулярная организация их определяется особенностями взаимодействия белковых молекул, так как им передается генетическая информация, необходимая для построения клеточных структур. По общепринятому мнению к межмолекулярным взаимодействиям, стабилизирующим биологические мембраны, относятся электростатические силы, силы Лондона — Вандер-Ваальса и гидрофобные взаимодействия. Показано, что наиболее важное значение в общей энергии межмолекулярных связей структурных и функциональных компонентов мембран принадлежит гидрофобным взаимодействиям (Гельмане сотр., 1972).
Исследования показали, что прочность межмолекулярных связей белков в протопластах клеток галловой ткани филлоксероустойчивых сортов всегда выше, чем у неустойчивых сортов винограда, о чем можно судить по вымываемости белков. Чем меньше их выделяется в буферный раствор, тем прочнее они связаны в мембранах (Соколовская, 1969; Зотов, 1976).
Под действием буферного раствора неионогенного детергента тритон Х-100 из пораженных тканей корней филлоксероустойчивых сортов винограда белка выделяется в 2—2,5 раза меньше, чем из неустойчивых сортов. Большая доля его приходится на непосредственное действие детергента тритон Х-100, который действует на гидрофобные межмолекулярные связи мембран клеточных структур (табл. 2).
Та же закономерность наблюдается при исследовании ткани листьев, пораженных филлоксерой. В этом случае различия между устойчивыми и неустойчивыми сортами еще больше выражены (табл. 3).
Таблица 3
Процент выделившегося белка (по азоту) из 10-дневных, пораженных филлоксерой листьев винограда в трис-цитратном буфере pH 8,6 без детергента и с детергентом тритоном Х-100 (mg/g сырой ткани)
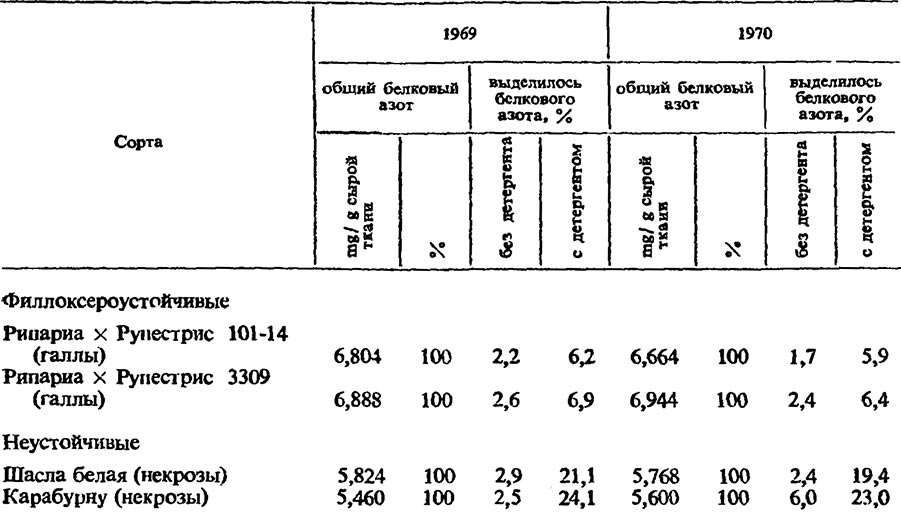
Следовательно, характер связи белка в биомембранах филлоксероустойчивых сортов обусловливает большую прочность клеточных структур, которые в условиях неблагоприятного воздействия филлоксеры продолжают выполнять свои функции. У неустойчивых сортов винограда под воздействием слюны филлоксеры в протопласте клеток наступают деструктивные изменения вследствие слабых связей белка с другими компонентами биомембран клеточных органоидов. Такие различия между устойчивыми и неустойчивыми к филлоксере сортами винограда определяют направленность биохимических процессов, развивающихся в галловой ткани.
В развитии защитной реакции решающее значение имеет запасание энергии. Образование митохондрий вблизи ядерной оболочки и накопление их в большом количестве в клетках галловой ткани филлоксероустойчивого сорта винограда говорят о том, что в пораженной ткани повышается энергетический обмен. Было также известно, и теперь это уже доказано, что в клеточных ядрах происходит окислительное фосфорилирование (А11еf rеу, Мirsку, 1957; Бетел, 1972; Рубин, Логинова, 1973). Дифференциальным центрифугированием из галловой ткани мочковатых корней винограда выделили фракцию ядер и прилегающих к ним мелких гранул, среди которых значительную часть составляют митохондрии. В этой фракции было изучено окислительное фосфорилирование (Зотов, 1964). Из данных явствует, что в филлоксерных галлах устойчивого сорта винограда окисление сопряжено с фосфорилированием, в то время как у неустойчивого сорта наблюдается разобщение этих процессов и окислительное фосфорилирование оказывается полностью подавленным.
Содержание нуклеиновых кислот в здоровых и поврежденных филлоксерой тканях листьев винограда (mg/100 cm листовой пластинки) (по Сторожук, 1970)
Таким образом, у устойчивых к филлоксере форм винограда освобождающаяся энергия дыхания аккумулируется в форме макроэргических фосфатных связей, которые используются для различных форм защитных реакций, прежде всего, для усиления синтетического направления обмена и подавления гидролизующего действия ферментов.
Обмен нуклеиновых кислот и белков является центральным процессом жизнедеятельности клеток и, безусловно, лежит в основе всех изменений, наступающих в пораженных филлоксерой тканях винограда.
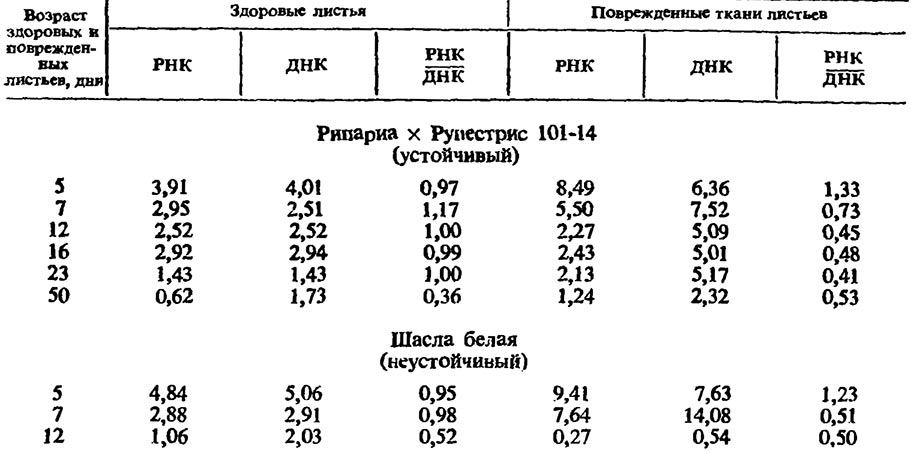
Исследованиями ряда авторов было показано, что в листьях и корнях винограда филлоксера вызывает характерные для устойчивых и неустойчивых сортов нарушения обмена нуклеиновых кислот (Светлякова с сотр., 1966, Зотов с сотр., 1966; Сторожук, 1970). При образовании некротических пятен на листьях неустойчивого к филлоксере сорта Шасла вспышка усиления биосинтеза нуклеиновых кислот непродолжительна. Уже на 12-й день после укола филлоксеры содержание РНК и ДНК в них снижается в четыре раза по сравнению со здоровыми листьями этого возраста (табл. 4).
Такое низкое содержание нуклеиновых кислот является критическим. Ткань в это время гибнет.
На листьях филлоксероустойчивого сорта образуются нормальные галлы, в которых на двенадцатый день после укола тли содержание РНК близко к норме, а количество ДНК возрастает в два раза по сравнению со здоровыми листьями. На 50-й день нуклеиновых кислот содержится в два раза больше, чем в здоровых листьях этого же возраста (табл. 4).
Аналогичная закономерность наблюдается и в корневых филлоксерных галлах устойчивого сорта. В них происходит постепенное накопление нуклеиновых кислот (табл. 5), они остаются живыми в течение всего вегетационного периода и перезимовывают. На той же табл. 5 видно, что в корневых галлах неустойчивого сорта РНК не накапливается. В 4—5-й декаде после укола филлоксеры корневые галлы гибнут.
Корневые галлы образуются, в основном, в результате увеличения размера клеток.
Таблица 5
Содержание нуклеиновых кислот в здоровых мочковатых корнях и корневых галлах винограда (mg/g сырой ткани) (по Сторожук, 1970)

Поэтому расчеты РНК, ДНК и белков сделаны на единицу сырой ткани. При образовании же листовых галлов лист утолщается за счет числа клеток, нарастающих вследствие деления клеток при развитии галла. В связи с этим расчеты веществ сделаны на площадь листовой пластинки.
Изменения, вызванные филлоксерой в белковом обмене в поврежденных ее тканях, протекают в такой же закономерности, как в нуклеиновом. В корневых галлах филлоксероустойчивого сорта винограда Рипариа х Рупестрис 101—14 происходит постепенное накопление белков с 2,42 mg/g на 15-й день до 3,36 mg/g на 40-й день. У неустойчивого сорта Шасла белая после незначительного увеличения на 15-й день (2,05 mg/g) количественное их содержание на 40-й день уменьшается более чем в два раза (0,62 mg/g) по сравнению со здоровыми корнями. В пораженных филлоксерой тканях листьев устойчивых сортов Рипариа х Рупестрис 101-14 и 3309 количество белков на 10-й день также резко возрастает (с 12,8 до 34,1 и с 12,6 до 38,3 mg/100 cm2), а у неустойчивых соответственно снижается с 8,9 до 7,5 (Шасла белая) и с 11,2 до 10,2 mg/100 cm2 (Карабурну).
Аминокислотный состав белков в тканях винограда, пораженных филлоксерой, изменяется. По данным Соколовской (1966), в белках листовых и корневых галлов устойчивого сорта Рипариа х Рупестрис 101-14 найдены те же аминокислоты, что и в здоровых мочковатых корнях, но в других соотношениях. Содержание цистина и оксипролина немного снижается, триптофан остается без изменения, а содержание остальных аминокислот увеличивается в 2—4,5 раза. В этом отношении особенно выделяется метионин и гистидин. Такие же изменения происходят в белках листовых галлов. Изменения соотношения аминокислот этих белков свидетельствуют о том, что их состав в количественном отношении изменяется. По всей вероятности, происходит биосинтез специфических белков, свойственных только пораженным филлоксерой тканям. У неустойчивого к филлоксере сорта Шасла в тканях листьев и корней, пораженных филлоксерой, соотношение аминокислот в белках не изменяется. В белках корневых галлов этого сорта метионин, триптофан, цистеин и оксипролин не обнаружены, а в пораженных листьях отсутствует триптофан и метионин.
Прекращение включения некоторых аминокислот в белки больных тканей неустойчивого к филлоксере сорта показывает, что на определенном этапе заболевания биосинтез целого ряда белков не происходит, в том числе и каталитических, а это приводит к глубокому нарушению общего метаболизма клеток и отмиранию тканей.
Примером белка-фермента, биосинтез которого прекращается, может быть цитохромоксидаза. Арциховская и Денисова (1966) наблюдали, что в галлах неустойчивого сорта Шасла активность цитохромоксидазы резко падает, а перед началом отмирания пораженной ткани этот фермент в галлах совсем не удается обнаружить. В то же время в галловой ткани филлоксероустойчивого сорта Рипариа х Рупестрис 101-14 активность цитохромоксидазы возрастает в 2,5—3 раза. Такая же закономерность наблюдается в отношении ферментов полифенолоксидаза и пероксидаза (Зотов, 1964; Арциховская, Денисова, 1966).
Следует отметить, что в клетках галловой ткани накапливается крахмал, причем в корневых галлах филлоксероустойчивых сортов более чем в 2—3 раза больше, чем у неустойчивых (Зотов, 1955).
Накопленный экспериментальный материал позволяет сделать следующее заключение.
У филлоксероустойчивого сорта опухолевые ткани захватывают лишь верхние слои коры. Усиление синтеза РНК и белков, повышение общего уровня энергетического обмена обеспечивает развитие синтетического направления обмена и жизнь клеток в тканях, пораженных филлоксерой. Это благоприятствует возникновению еще под живой опухолью феллогена и дальнейшему формированию защитной раневой перидермы, которая отслаивает на корнях небольшие участки отмершей опухолевой ткани. В результате в тканях, находящихся под раневой перидермой, обмен продолжается, корни остаются живыми и заболевание оканчивается самоизлечением. Это происходит как на мочковатых корнях первичного строения, так и на одревесневших корнях. Эти же процессы на листьях устойчивого сорта в местах сосания филлоксеры приводят к образованию нормальных галлов, ткани которых остаются живыми до осеннего листопада.
У сорта винограда, неустойчивого к филлоксере, образование опухолей протекает в глубоких слоях тканей корней, захватывающих кору и часть древесины. Биосинтез РНК и белков затухает. Общий уровень энергетического обмена резко снижается и гидролитические процессы усиливаются. Вполне понятно, что в таких условиях не может возникать образовательная ткань — феллоген, раневая перидерма не формируется и на корнях после отмирания опухолевых клеток образуются большие открытые раны, которые загнивают. Эти же процессы на листьях неустойчивого сорта в местах сосания филлоксеры приводят к возникновению некрозов.
Изложенное выше свидетельствует о том, что ответная реакция на поражение филлоксерой, возникающая в тканях корней и листьев, характеризуется тождественными биохимическими процессами, характер которых зависит от степени устойчивости сорта, а морфологические признаки пораженных филлоксерой листьев отражают процессы, происходящие в пораженных тканях и также,как пораженные корни, могут служить для определения филлоксероустойчивости сортов и форм винограда.
На рис. 2 представлена анатомическая картина описанных процессов на мочковатых корешках первичного строения, одревесневших корнях и листьях.
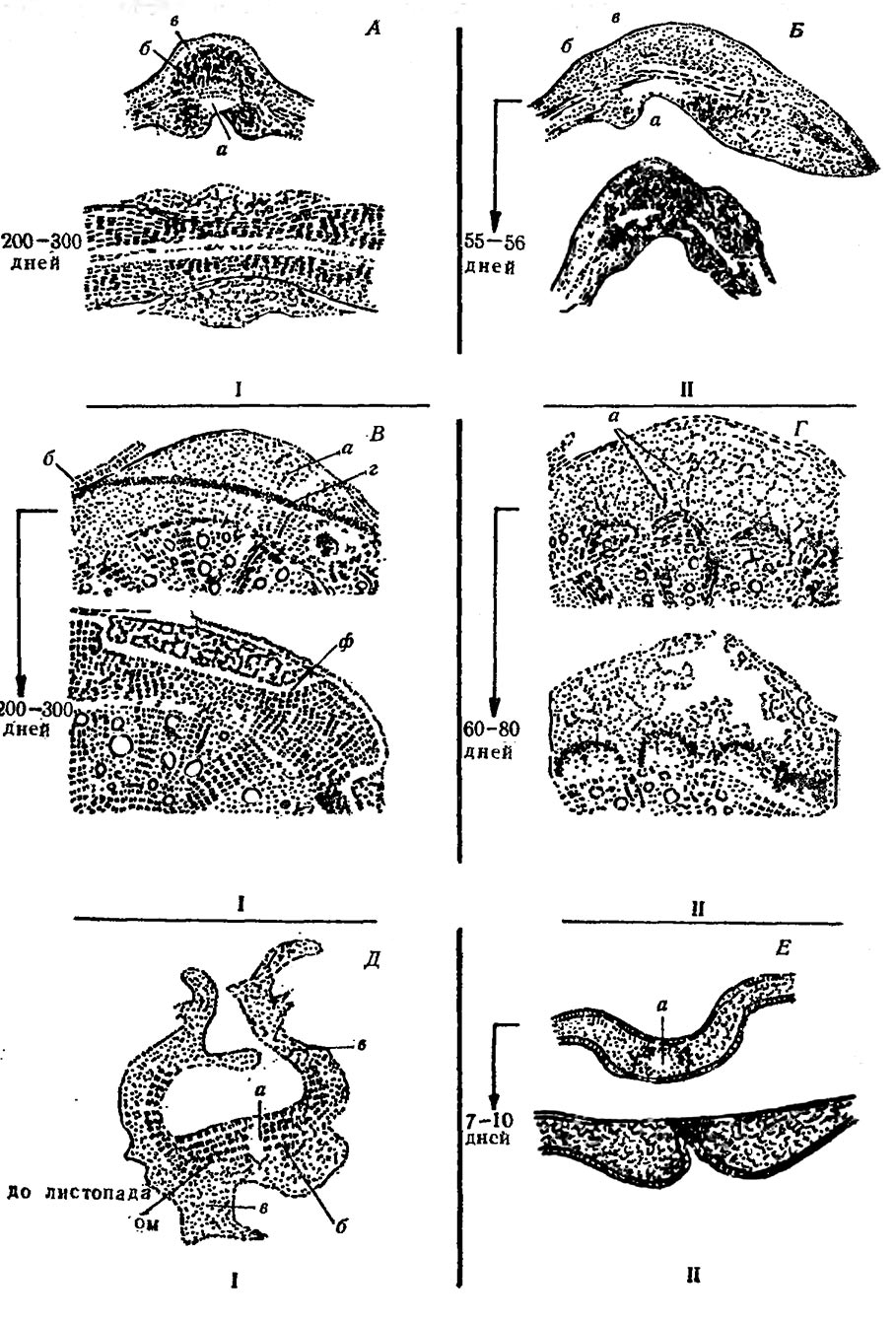
Рис. 2. Анатомические изменения в корнях и листьях винограда под влиянием филлоксеры Рипария х Рупестрис 101-14 — устойчивый, Шасла — неустойчивый
А, Б— корневые галлы; I— отслаивание, II— гибель ткани; В, Г— одревесневшие корни: I- отслаивание, II — гибель ткани; Д, Е— листья: I — образование листового галла, II — гибель ткани, образование некротического пятна
а — зона гидролиза (место питания филлоксеры); б — зона синтеза; в— зона растяжения; г — слой клеток, богатых РНК; ф — раневая перидерма; ом — остаточная меристема
Познание физиологической сущности устойчивости винограда к филлоксере необходимо прежде всего для того, чтобы составить ясное представление о том, какими свойствами должны обладать филлоксероустойчивые сорта винограда, и на этой основе определить устойчивость селекционных форм.
Провокационный фон и отбор на основе искусственного заражения и гибели неустойчивых растений от вредителей оставляют много неясных вопросов, особенно при размножении выделенных форм. На развитие филлоксеры влияет механический состав почвы и ее физические свойства (Принц, Иванов, 1948; Казас с сотр., 1960). Агротехнические условия влияют на сопротивляемость филлоксере кустов неустойчивых сортов винограда (Мордвилко, 1925; Зотов, 1955; Казас с сотр., 1960; Михайлов, 1960). Все, что снижает синтетическую функцию в корнях, ускоряет гибель кустов, зараженных филлоксерой. Мощные формировки, допускающие накопление на кустах многолетней древесины, небольшая нагрузка кустов урожаем, поливы, регулярное внесение удобрений, подкормки и пр. — все это повышает сопротивляемость кустов. Напротив, малые формировки без многолетней древесины, перегрузка кустов урожаем, частые дефициты влаги в летние месяцы, истощенная почва снижают сопротивляемость кустов винограда, зараженных филлоксерой. Время заражения также играет большую роль. При заражении в 3—5-летнем возрасте состояние явного угнетения может наступить на 10—15-й год. В этом случае нетрудно допустить ошибку, сделав преждевременный вывод.
В виноградарстве всегда ощущалась острая необходимость быстрых и точных методов определения филлоксероустойчивости сортов винограда, Фоэкс (1904) обратил внимание на различие анатомического строения корней устойчивых и неустойчивых к филлоксере сортов винограда. Фундаментальное исследование связи между филлоксероустойчивостью винограда и анатомическим строением корней провели Абесадзе с сотр, (1930) при изучении устойчивости грузинских сортов винограда. Они нашли, что для устойчивых сортов характерны толстые оболочки клеток, узость сердцевинных лучей, мелкоклеточность, небольшая сердцевина, тонкая кора. Корни неустойчивых сортов характеризуются рыхлым строением тканей, крупноклеточностью, широкими сердцевинными лучами и- толстой корой. Однако мочковатые корешки первичного строения по анатомическому строению не отличаются. Эти авторы установили, что у устойчивых сортов в местах поражения корней филлоксерой раневая перидерма образуется более интенсивно, чем у неустойчивых сортов. В результате проведенных исследований были выделены относительно устойчивые сорта — Чинури, Мцване, Ркацители — и сделан вывод, что среди грузинских сортов винограда необходимо вести клоновую селекцию на филлоксероустойчивость.
Вorner (1942) придает большое значение анатомическому строению корней в филлоксероустойчивости винограда.
Он утверждает, что особенности анатомии корней филлоксероустойчивых сортов способствуют отслаиванию поврежденных филлоксерой участков ткани. По этому принципу он разделил виды винограда по степени филлоксероустойчивости. Бернер считает, что у европейских сортов винограда могут происходить мутации в направлении повышения филлоксероустойчивости.
В. В. Зотов придает большое значение состоянию корневых галлов, образующихся на мочковатых корнях первичного строения. Автор установил, что корневые галлы неустойчивых сортов быстро отмирают, вследствие чего мочковатые корешки гибнут. Кусты остаются без всасывающих корней и страдают от недостатка влаги. С этого начинается угнетающее действие филлоксеры. У филлоксероустойчивых сортов после образования корневого галла мочка, приобретающая вторичное строение, продолжает нормально расти и остается нормальной толщины. При дальнейшем росте на ней может образоваться еще несколько корневых галлов подобно бусам, нанизанным на шнурок. Ветвление мочковатого корешка продолжается нормально. Корневые галлы покрываются опробковавшей тканью, остаются живыми в течение всего вегетационного периода и перезимовывают. Весной клетки, образующие опухоль, отслаиваются раневой перидермой, а корешок остается живым. На основе описанных различий предложен метод определения филлоксероустойчивости по способности корневых галлов перезимовывать и весной изолировать опухолевую ткань (Зотов, 1955). Важно заметить, что мочковатые корешки первичного строения по анатомическому строению не отличаются; тем не менее, в образовавшихся на них корневых галлах протекают, как это было описано выше, биохимические процессы различной направленности, которые у устойчивых сортов приводят к образованию защитной раневой перидермы, а у неустойчивых — к летальному исходу.
Соколовская (1961) предложила пятибалльную систему определения филлоксероустойчивости сортов винограда по анатомо-цитологическим и морфологическим признакам пораженных филлоксерой корней. Согласно этой методике для оценки филлоксероустойчивости изучают следующие изменения корней, вызванные филлоксерой: 1) морфологические признаки корневых галлов и характер ветвления мочковатых корешков, пораженных филлоксерой; 2) способность корневых галлов перезимовывать; 3) заполнение крахмалом клеток первичной коры корневых галлов; 4) состояние клеточных ядер и цитоплазматических гранул в клетках первичной коры сформировавшихся корневых галлов со стороны зоны роста корневого галла; 5) характеристика деятельности камбия и феллогена на многолетних корнях в местах поражения филлоксерой (интенсивность образования раневой перидермы). Каждому признаку дается оценка в пяти баллах; чем выше устойчивость, тем выше балл. Дана балльная оценка каждого признака, по которому ведется наблюдение, общий балл по сорту выводится путем вычисления средней со значением: от I до 2 баллов — неустойчивый, от 2 до 3,8 — относительно устойчивый, от 3,8 до 5 — устойчивый сорт, пригодный для разведения на своих корнях. Для исследования берут не менее 10 повторностей.
Эта методика удобна тем, что она дает возможность учесть соотношение типов поражения корней филлоксерой. Дело в том, что на корнях и листьях виноградного куста, особенно на промежуточных по устойчивости сортах, наблюдаются различного типа повреждения. На корнях одного и того же куста встречаются открытые раны и поврежденные участки, частично или полностью защищенные раневой перидермой. Листья на побегах куста также отличаются по типу ответной реакции на поражение филлоксерой.
Таблица 6
Варьирование количества нормальных галлов (в процентном содержании) на листьях отдельных побегов различных сортов винограда
Сорт и его характеристика | Варьирование от — до | Среднее по сорту |
Рипариа х Рупестрис 101-14, филлоксероустойчивый подвойный сорт | 95—99 | 97,5 |
Рииариа Глуар, филлоксероустойчивый вид | 91—99 | 97,9 |
Рупестрис Бринье, филлоксероустойчивый вид | 93—96 | 94,6 |
Корна нягра, относительно устойчивый европейский сорт Шасла белая, неустойчивый европейский сорт | 42—80 | 50,4 |
0—28 | 10,0 | |
Голубок, новый сорт, межвидовой гибрид | 23—82 | 54,6 |
Находка, новый сорт, межвидовой гибрид | 34—82 | 65,2 |
Терновый, новый сорт, межвидовой гибрид | 29—84 | 65,5 |
Могут образоваться некрозы, блюдца (признаки неустойчивости), стерильные и нормальные галлы (признаки устойчивости). Это явление следует объяснить гетероплазматическим состоянием тканей виноградного растения по степени филлоксероустойчивости. При определении филлоксероустойчивости сортов винограда и отборе устойчивых форм очень важно учитывать соотношение этих типов повреждения. Чем больше на корнях повреждений, защищенных раневой перидермой, и нормальных галлов на листьях, тем устойчивее куст (Зотов, 1976б).
Соотношение повреждения листьев на отдельных побегах на кусте варьирует по сорту в определенных пределах (Зотов, 1976б; табл. 6).
Изучение гибридной семьи Корна нягра X Датье де Сен Валье показало большую сопряженность между количеством нормальных галлов на листьях и полностью защищенных раневой перидермой филлоксерных повреждений на корнях, выражающуюся коэффициентом корреляции r=0,84±0,06 (Зотов, 1976а). Такая сопряженность, как это было показано выше, обусловливается тождеством биохимических процессов, развивающихся в тканях листьев и корней, пораженных филлоксерой, которые зависят от степени устойчивости сорта.
На этой основе Зотовым (1967) разработана методика определения филлоксероустойчивости сортов и селекционных форм винограда по морфофизиологическим показателям пораженных филлоксерой листьев. Сорта и формы, у которых процентное содержание образовавшихся нормальных галлов на листьях приближается к 70—80 %, относят к филлоксероустойчивым, пригодным для культуры на своих корнях; свыше 80 % — к высокоустойчивым сортам. Листья (верхушки) побегов изучаемых сортов винограда нужно искусственно заражать филлоксерой. Этот метод может быть использован не только для определения устойчивости сортов, но и при клоновой селекции винограда на филлоксероустойчивость, так как дает возможность отбирать лозы на кустах. Новому кусту дает начало черенок. Поэтому при клоновой селекции и при размножении новых сортов нужно отбирать лозы, обладающие положительной наследственностью, на листьях которых в подавляющем большинстве случаев образуются нормальные галлы. Именно такие лозы регенерируют филлоксероустойчивые корни.
В редких случаях на некоторых сортах американских видов местный биотип филлоксеры не индуцирует галлообразования на листьях, а при заражении другими биотипами происходит образование нормальных галлов. Конечно, существование и появление новых рас патогенов усложняет иммуноселекцию сельскохозяйственных растений. Но единичные случаи, когда на каком-либо сорте американского вида местный биотип филлоксеры не индуцирует образования нормальных галлов, не нарушают общей закономерности, заключающейся в том, что лозы, образующие на листьях нормальные галлы, регенерируют филлоксероустойчивые корни. Важно отбирать селекционные формы, на листьях которых в подавляющем большинстве случаев развиваются нормальные галлы, и при вегетативном размножении отбирать по этому признаку лучшие лозы путем повторяющегося отбора.
Кискин (1965) предложил стобалльную оценку филлоксероустойчивоcти по 1) анатомическим признакам здоровых корней; 2) опухолеобразующей способности корней при поражении филлоксерой и 3) интенсивности образования раневой перидермы на корнях в местах поражения.
Различий в анатомическом строении корней первичного строения Кискин не нашел и все свои исследования проводил только на корнях вторичного строения, преимущественно в возрасте 2—4-х лет. Анатомическое строение корней изучается по 23 признакам, опухолеобразовательная способность при поражении корней филлоксерой — по 16 и интенсивность образования раневой перидермы — по 20. Каждый признак оценивается баллом. Чем большую роль играет признак в устойчивости корней к филлоксере, тем выше балл. Например, самые высокие баллы имеются тогда, когда опухоли захватывают лишь кору (более 60% случаев); камбий не прекращает работу; опухоли мелкие, глубиной 0,5 mm, составляют более 60%; опухоли отделены сплошным, хорошо развитым и пропитанным суберином слоем раневой перидермы (более 50% случаев); раневая пробка образуется быстро, происходит регенерация корней и др. Филлоксероустойчивость характеризуется по сумме набранных баллов: до 50 — неустойчивые, от 50 до 80 — среднеустойчивые, от 80 до 100 — филлоксероустойчивые сорта. По методике Π. X. Кискина работу проводят на сформированных плодоносящих кустах.
Недов(1975) считает, что корни многих гибридных и европейских сортов после поражения филлоксерой гниют с неодинаковой интенсивностью вследствие различия их устойчивости к гнилостным микроорганизмам. У некоторых сортов гниение протекает настолько медленно, что кусты долгое время могут находиться в удовлетворительном состоянии в условиях заражения филлоксерой. Такие сорта Π. Н. Недов относит к относительно устойчивым (толерантными) к филлоксере. Степень устойчивости сортов по методике Π. Н. Недова определяется глубиной гниения тканей корней после поражения филлоксерой. Для этих целей в грунт высаживают двух — трехглазковые черенки испытываемых сортов. Когда корни примут вторичное строение и одревеснеют, их заражают филлоксерой, вносят в почву загнивший ячмень с целью увеличения инфекционного запаса гнилостных микроорганизмов. Когда корни загнивают, производят анализы на степень развития в них гнилостных процессов.
Механически поврежденные корни неустойчивых сортов винограда не загнивают. Летом на корнях неустойчивого к филлоксере сорта Шасла делали надрезы ножом в коре одревесневших корней и уколы в зоне растяжения на мочковатых корешках первичного строения, чтобы создать доступ микроорганизмов к тканям. Оказалось, что в условиях контакта с землей места ранения покрываются защитной пробкой (Зотов, Маречек). Пораженные же филлоксерой корни Шасла погибают.
Воubals (1966) предложил ускоренный метод определения филлоксероустойчивости сортов винограда в теплице. Укороченные черенки испытываемых сортов винограда высаживают в тепличных стеллажах и после того, как корни разовьются, их заражают филлоксерой. Когда ответная реакция корней на поражение филлоксерой явно выражена, производят оценку на степень повреждения, которую Д. Бубаль разделяет на четыре класса. Класс 0 — на всех корнях туберозитеты (опухоли) не образуются. Класс I — на всех корнях образуются туберозитеты, но они немногочисленны. Ткань туберозитетов бывает некротизированной. Во всех случаях этот некроз локализован в корне и не достигает луба. Класс II Бубаль разделяет на два случая: а) туберозитетов образуется мало, но некроз ткани проникает очень глубоко в корень и частично поражает флоэму; б) туберозитеты располагаются на корне в цепочку, с более или менее глубокими некрозами, но корни не так разрушаются, как в третьем классе. Класс III — туберозитеты многочисленны, больших размеров и практически полностью разрушаются. Распад достигает центра корня. У наиболее сильно пораженных растений распадаются все корни. Сорта винограда, относящиеся к классам О и I, являются вполне филлоксероустойчивыми. Таким образом, в настоящее время селекционеры располагают методами определения филлоксероустойчивости сортов винограда и имеют возможность с большой точностью вести отбор устойчивых форм и их размножение на основе улучшающегося, повторяющегося отбора.