Мильдью — самая распространенная и наиболее опасная болезнь винограда. Поражает листья, соцветия, ягоды и зеленые побеги. Во влажные годы мильдью уничтожает значительную часть винограда. По данным Вердеревского с сотр. (1975) ежегодно затраты на борьбу с мильдью в целом по СССР выражаются в сумме около 30 млн. рублей, а потери урожая винограда от этого заболевания составляют 10—15%. Вред, причиняемый мильдью, не ограничивается только снижением урожая. Вследствие потери листьев виноградная лоза плохо вызревает и зимой легко повреждается морозами, а также снижается ценность ее в качестве посадочного материала.
Возбудитель мильдью — гриб Plasmopara viticola Berl. et de Toni — относится к класу Phycomyceteae (низшие грибы) семейства Peronosporaceae. Облигатный паразит,так как на всех стадиях онтогенеза ведет паразитический образ жизни только в живых тканях растения и в сапрофитных условиях не развивается. Мицелий гриба одноклеточный, многоядерный, развивается в межклетниках мезофилла листа, извлекает пищу при помощи гаусторий.
Органами бесполого размножения являются зооспорангии, которые при прорастании образуют двухжгутиковые зооспоры. Зимующей стадией гриба являются ооспоры, образующиеся в межклетниках тканей растения-хозяина в результате полового процесса. Биология развития гриба довольно полно освещена многими авторами (Принц, 1952, 1962; Вердеревский, 1947, 1958; Вердеревский, Войтович, 1970; Костюк, 1949; Липецкая, Рузаев, 1958; Макрушина, 1967; Горленко, 1968; Найденова, 1969).
Изучению вопроса о существовании физиологических рас Pl. viticola посвящены исследования ряда авторов. Так, работы Boubals (1957, 1959), Grunzel (1961), Рафаилэ (1965), Вердеревского с сотр. (1969) показали отсутствие физиологических рас в составе вида Pl. viticola, отличающихся специализацией к отдельным видам и сортам винограда и своей вирулентностью. Однако Santilli (1957) указывает на существование в условиях США двух различных физиологических рас Pl. viticola — западная и восточная. В пределах СССР, Китая по морфологическим признакам выделены несколько видов возбудителя мильдью,например, амурский (Проценко, 1946), среднеазиатский (Головина, 1955), китайский (Горленко, 1969).
Исследования Вердеревского с сотр. (1969) по изучению патогенности штаммов Pl. viticola различного происхождения (Молдавия, Крым, Сочи, Дальний Восток) не выявили, однако, каких-либо существенных различий между испытанными штаммами.
Для развития мильдью необходимо определенное сочетание внешних условий, из которых важнейшими являются температура и влажность. Развитие заболевания с момента заражения до появления желтых "маслянистых" пятен на листьях носит название скрытого или инкубационного периода. Его продолжительность зависит, главным образом, от среднесуточной температуры. Нижним порогом температуры, необходимым для развития грибницы мильдью в тканях листьев, является +8°, а верхним порогом +32° (Липецкая, 1937; Шатский, 1939). Мюллер и Рабанус (1923) установили зависимость продолжительности инкубационного периода от среднесуточной температуры, где эта зависимость изображена в виде кривой, параметры которой представлены в табл. 7 (цит. по Костику, 1949).
Самой благоприятной температурой воздуха для развития мильдью является 22—24°. Кривая Мюллера позволяет ориентировочно определять сроки химической обработки виноградников.
Развитие мицелия после внедрения зооспор через устьица в листьях винограда зависит от степени устойчивости сорта. Все культивируемые высококачественные европейские сорта принято считать неустойчивыми, хотя в пределах вида (Vitis vinifera) имеются сорта, обладающие повышенной сопротивляемостью к мильдью.
Устойчивостью обладают дикие американские виды винограда, на листьях которых при поражении мильдью возникают мелкие точечные некрозы диаметром 1—2 mm2.Только при наличии очень повышенной влажности по краям некротических пятен могут образоваться единичные, слаборазвитые конидиеносцы. На листьях американо-европейских гибридов, обладающих промежуточной устойчивостью, мильдью образует пятна более крупных размеров, пораженная ткань в центральной части быстро отмирает. Спороношение гриба наблюдается лишь по краям пятна. В листьях же восприимчивых культурных сортов V.vinifera мицелий гриба быстро развивается,захватывая большие участки листовой пластинки, и обильно спороносит.
Естественный отбор способствовал созданию форм винограда, устойчивых к мильдью, т. е. созданию специфического иммунитета, который обусловливается, прежде всего, характером реакции живой ткани на воздействие патогена.
Таблица 7
Продолжительность инкубационных периосов развития мильдью
Температура воздуха, °C | Продолжительность инкубационного периода, сутки | Температура воздуха, °С | Продолжительность инкубационного периода, сутки |
12 | 13 | 21 | 4,5 |
13 | 11 | 22 | 4 |
14 | 9,5 | 23 | 4 |
15 | 8,5 | 24 | 4 |
16 | 7 | 25 | 4,5 |
17 | 6,5 | 26 | 4,5 |
18 | 5,5 | 27 | 5 |
19 | 5 | 28 | 6 |
20 | 4,5 | 29 | 8 |
Европейские сорта, находившиеся за пределами первоначального распространения возбудителя мильдью, оказались неустойчивыми к этому патогену.
Согласно Негрулю (1959), все роды семейства Vitaceae и подрод Muscadinia рода Vitis (V. rotundifolia и V. Munsoniana) обладают пассивным иммунитетом к мильдью. Только подрод Euvitis рода Vitis включает в себя виды винограда от высокоустойчивых до высоковосприимчивых (V. vinifera). Это подтвердил Воubals (1959) своими исследованиями на большом экспериментальном материале. Согласно автору, роды семейства Vitaceae по устойчивости к мильдью располагаются в следующем порядке: Tetrastigma → Cissus → Parthenocissus → Ampelopsis → Vitis. В эволюционном развитии американских видов винограда, как указывают Вердеревский с сотр. (1975), осуществлялись ступени естественного образования комплексно устойчивых видов к мильдью, оидиуму, филлоксере.
Для оценки степени мильдью устойчивости винограда разработаны шкалы, основанные на особенностях развития болезни на листьях, на размерах пораженного участка листовой пластинки, интенсивности спорообразования гриба и скорости отмирания пораженных тканей листьев (Sеeligеr, 1926; Arnoud, 1931; Нusfeld , 1933; Зотов, 1935; Вoubals , 1959; Шапа , 1965; Найденова, 1962).
Arnoud (1931) предложил обширную 20-балловую шкалу определения устойчивости для видов и сортов рода Vitis. Шкала очень громоздкая и неудобная для применения.
Шапа (1965), упрощая шкалу Арно, выделил из коллекционного фонда Молдавского научно-исследовательского института садоводства и виноградарства 10 групп по мильдью устойчивости, способных служить эталоном при оценке новых форм винограда.
Seeliger (1926), Husfeld (1933), Зотов (1935) предложили 5 типов поражения, основанные на интенсивности спороношения и размерах пораженных тканей, а также скорости их отмирания.
Вoubals (1959) распределяет виды рода Vitis по степени устойчивости на пять классов: в I класс автор относит наиболее высокоустойчивые виды: V. aestivales, V. rubra.V. riparia; класс II — V. cordifolia, V. candicans, V. rupestris; класс III — V. Berlandieri, V. monticola; класс IV — V. lincicumii, класс V — наиболее восприимчивые виды — V. vinifera, V. californica, V. arisonica.
Кроме того, автор вводит в шкалу нулевую группу, куда относит V. rotun- difolia и V. Wilsonae, обладающие пассивным иммунитетом.
Работы Д. Бубаля имеют большое значение для селекционеров и генетиков. Однако в шкале автор не дает градаций по устойчивости для сортов V. vinifera. Найденова (1962) дает оценку мильдью устойчивости селекционного материала на основе шкалы Гусфельда, дополненной показателем интенсивности заражения (количество пораженных листьев на кусте). Кроме того, учитывается поражаемость генеративных органов. Автор приводит формулу для определения степени поражения куста.
С целью ускорения оценки селекционного материала Штин (1969) и Щербатюк (1964) предлагают метод искусственного заражения изолированных листьев в пробирках или специальных камерах.
Все существующие шкалы могут быть использованы для оценки мильдью устойчивости винограда. При этом необходимо учитывать величину инфекционной нагрузки и варьирование внешних условий. Как известно, физиологическое состояние растения изменяется как от климатических факторов (температура, влажность), так и на протяжении вегетационного периода.
В настоящее время в Советском Союзе селекционеры пользуются в основном 5-балльной шкалой, где баллом 1 оцениваются наиболее устойчивые виды и сорта, а баллом 5 — более восприимчивые сорта и формы винограда. Например, Гузун с сотр. (1975) приводят следующие параметры для оценки мильдью устойчивости винограда по 5-балльной шкале: балл 1 — отдельные некрозные точки, пушка нет; балл 2 — редкие некрозные пятна, пушка нет;
балл 3 — наличие некрозных больших пятен во всех листьях, имеется пушок; балл 4 — все листья поражены, 50—60% листьев опало; балл 5 — все побеги без единого листа.
Оценка устойчивости к мильдью по степени поражаемости листьев в условиях достаточного провокационного фона является основным критерием при изучении селекционного материала. Оценка проводится в течение нескольких лет.
В целях совершенствования и ускорения методов селекции и генетики большое значение имеет познание биохимической сущности иммунитета виноградных лоз к мильдью.
Изучению причин и сущности мильдью устойчивости винограда посвящен ряд исследований. Еще Аверна-Сакка (1910), Комес (1916), Нистеракис (1943) объясняли мильдью устойчивость различиями осмотического давления в клетках и кислотностью клеточного сока; различиями в содержании антоцианов, аминокислот, крахмала, сахаров, азота, калия (цит. по Вавилову, 1964). Arens (1929), Нusfeld (1933), Мержаниан (1935) связывали устойчивость винограда с морфолого-анатомическими особенностями листьев, такими, как количество устьиц, характер строения замыкающих клеток,плотное или рыхлое строение мезофилла листа, восковой налет и опушенность листьев. Установлены определенные различия между устойчивыми и восприимчивыми сортами по этим показателям.
Воubals (1959) указал на второстепенную роль анатомо-морфологических особенностей в устойчивости и высказал мысль о том, что в основе иммунитета винограда к мильдью лежат внутренние факторы, препятствующие развитию патогена.
Piоth (1957) выдвинула гипотезу, согласно которой восприимчивость винограда к мильдью обусловлена большим содержанием в листьях неустойчивых сортов солей щавелевой кислоты и индольных соединений. Однако в исследованиях Воubals (1959), Плисс (1968) наличие такой закономерности не подтвердилось.
Имеются исследования органических компонентов листьев винограда и спор гриба с целью определения пищевой полноценности виноградного растения к данному патогену (Sсuf1airе , 1967; Steward, Bidwell, 1962; Плисс, 1968а). По данным Р. Скюфлера, споры мильдью богаты сахарами, органическими кислотами, аминокислотами. Им обнаружено 4 сахара: глюкоза, фруктоза, сахароза, ксилоза; 22 аминокислоты. В. М. Плисс выделил из спор гриба 19 аминокислот. Из аминокислот больше всех содержатся глютамин, аспарагиновая кислота, триптофан, серин, треонин. Однако Р. Скюфлер пришел к выводу, что сравнительное изучение данных компонентов не позволяет определить пищевых потребностей патогена.
В настоящее время широкое признание и развитие получило учение об иммунитете культурных растений к болезням и вредителям, развиваемое Б. А. Рубиным и его школой, согласно которому иммунитет растительной клетки к патогенным организмам определяется, прежде всего, характером тех сдвигов в обмене веществ, которые наступают при взаимодействии растения-хозяина и паразита (Рубин с сотр., 1947, 1975; Рубин, Арциховская, 1960).
Большой вклад внесли исследования Л. В. Метлицкого и его сотрудников для познания молекулярных механизмов сверхчувствительности, а именно изучение химической структуры и биологического действия тех веществ, которые возникают в инфицированной клетке (Метлицкий с сотр., 1969; Метлицкий, Озерецковская, 1968, 1973; Метлицкий, 1976).
Важные исследования проведены Д. Д. Вердеревским и его сотрудниками по изучению причин мильдью устойчивости винограда (Вердеревский , 1958; Вердеревский с сотр., 1964; Шапа , 1959, 1965; Весьминьш, 1964, 1966; Найденова, Весьминьш, 1968; Найденова, 1969). Авторы наблюдали зависимость между степенью устойчивости и фитонцидной активностью клеточного сока листьев, которая связана с физиологическими процессами — активностью окислительных ферментов и интенсивностью дыхания. При этом фитонцидная активность инфицированных тканей выше, чем интактных. Повышенная фитонцидная активность устойчивых сортов рассматривается как защитная реакция растений против мильдью.
Какие вещества образуются в пораженных тканях, какие из них вызывают отмирание тканей и патогена?
Имеются разные точки зрения. Одни исследователи, как пишет Метлицкий (1976), отводят решающую роль фитоалексинам, другие рассматривают в качестве основного защитного механизма имеющуюся в растениях систему полифенол-полифенолоксидаза, под действием которой накапливаются продукты окисления полифенолов.
Пораженная ткань — очень сложная система. При этом действует сложная цепь различных взаимообусловленных процессов. Вот почему отдать предпочтение той или иной защитной системе весьма трудно.
Р. Ш. Гадиевым проведены исследования по изучению продуктов метаболизма здоровых и пораженных мильдью листьев винограда.
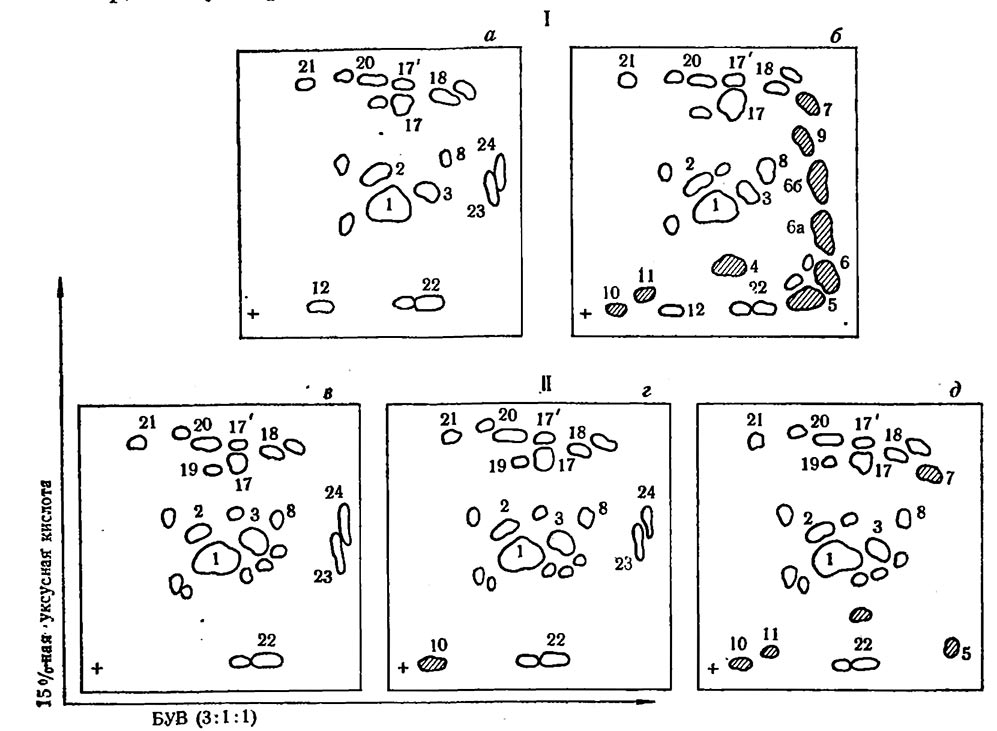
Рис. 3. Схема двухмерной хроматографии на бумаге этанольных экстрактов здоровых и пораженных мильдью листьев винограда
I — здоровый (а) и пораженный (б) лист Рипариа X Рупестрис 101-14 (устойчивый); Карабурну, здоровый лист (в), на 5—6-й день после поражения (г), на 9—10-й день после поражения (д)
вещества, отсутствующие в здоровых листьях. Показана высокая токсичность их к зооспорангиям мильдью. В метанольных экстрактах некротических тканей Рипариа х Рупестрис 101-14 при двухмерной хроматографии (I./1-бутанол—уксусная кислота — вода (БУВ) 3:1:1; II. 15 %-ная уксусная кислота) обнаружено 9 вновь образующихся веществ в результате искаженного обмена. Уже на 3—4-й день после заражения они накапливались в большом количестве, что приводит к образованию некроза (Гадиев, 1969; Гадиев, Зотов, 1971, 1971а).
Пораженные ткани восприимчивого культурного сорта Карабурну взяты в три срока: на 5-й день (до спороношения, 6—8-й день (период спороношения) и 9—10-й день — период отмирания тканей листьев. Образование защитных веществ наблюдается лишь на поздних этапах болезни. Они содержатся в очень незначительном количестве и не влияют на ход патогенеза (рис. 3).
Таблица 8
Активность полифенолоксидазы и пероксидазы в листьях винограда (выражено пурпурогаллиновым числом — n.ч.)
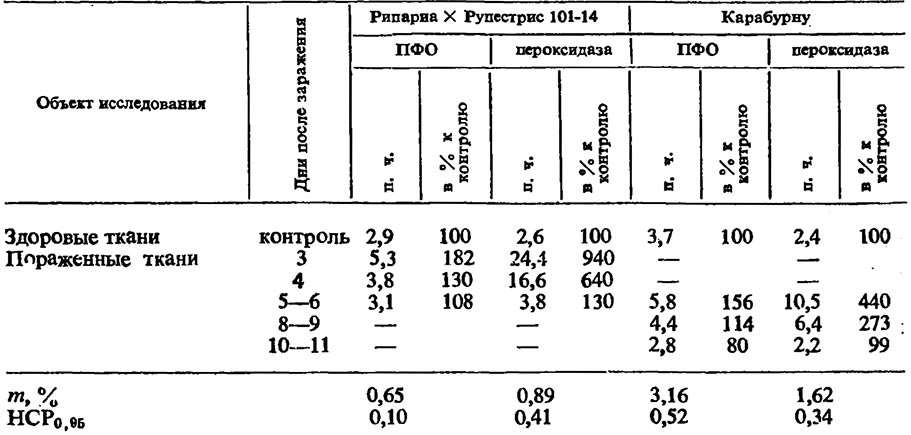
Таблица 9
Изменение интенсивности, дыхания листьев винограда при поражении мильдью (в ст3O2) поглощенного 1 g сырых тканей за 1 h)

Вещества, образующиеся в инфицированных тканях листьев,представлены на рис. 3 в заштрихованном виде и обозначены цифрами 4, 5,6,6а, 66,7, 9, 10, 11. Все они в УФ-свете (254 nm) флюоресцируют синим и голубым цветом различной интенсивности. По своей химической природе их можно отнести к липидам и фенолам. Вещества 4, 5, 7, 9 дают с 0,05 %-ным спиртовым раствором родамина лиловую окраску (в УФ-свете), типичную для фосфолипидов (Смирнов с сотр., 1961), а вещества 6, 6а, 66 реагируют с диазотированной сульфаниловой кислотой (ДСК), проявляя желтую и желтооранжевую окраску, характерную для фенольных соединений. Кроме того, с ДСК реагируют также вещества 7, 9 с желтым цветом.
На 35 сортах винограда, отличающихся по устойчивости к мильдью, выявлена прямая корреляция между интенсивностью и скоростью образования антибиотических веществ в пораженных тканях листьев и степенью устойчивости, о чем можно судить по количеству пятен на хроматограммах и интенсивности свечения их в УФ-свете.
Значительные различия в больных тканях устойчивого и восприимчивого сорта обнаружены также в составе фенолкарбоновых кислот. В здоровых листьях Рипариа х Рупестрис 101-14 и Карабурну найдены до 9 кислот: два изомера хлорогеновой кислоты (17), кофейная (8), феруловая (18), хинная (21) и другие неидентифицированные кислоты. При инфекции устойчивый сорт отличается значительным увеличением содержания хлорогеновой и кофейной кислот. Известно, что эти кислоты — активные участники различных биохимических процессов. Они являются основным субстратом в деятельности полифенолоксидазы (Метлицкий с сотр., 1972).
Заболевание мильдью винограда сопровождается значительными нарушениями ряда физиологических процессов. Большие сдвиги наступают в интенсивности дыхательного газообмена и активности окислительных ферментов. По данным Гадиева (1971) в пораженных тканях резко повышается активность пероксидазы: у устойчивого сорта на 3-й день после заражения активность фермента возрастает более чем в 9 раз; у восприимчивого — на 5-й день в 4, 5 раза (табл. 8). Активность полифенолоксидазы в пораженных тканях обоих сортов повышается в 1,8—2 раза.
Деятельность пероксидазы и полифенолоксидазы тесно связана с дыхательным газообменом. В пораженных тканях обоих сортов интенсивность дыхания возрастает в 1,8—2 раза больше, чем в интактных тканях (табл. 9). При этом необходимо учесть, что в пораженных тканях восприимчивого сорта повышение интенсивности дыхания частично связано с дыханием самого патогена,мицелий которого на 5—6-й день имеется в большом количестве в мезофилле листа.
Интересные данные выявил Весьминьш (1969) при изучении суточного хода изменения активности пероксидазы, полифенолоксидазы и фитонцидной активности пораженных мильдью листьев винограда, которые позволяют объяснить тот факт, что в ночное время, в период снижения жизнедеятельности клеток листьев, создаются условия, благоприятные для заражения и развития мильдью. С этим связано и выработанное в процессе эволюции свойство образования спороношения гриба в ночные часы.
Если рассмотреть качественный состав экстрактов листьев устойчивого и восприимчивого сортов до инфекции путем хроматографии на бумаге, у Рипариа х Рупестрис 101-14 можно обнаружить до 19, а у Карабурну — до 23 индивидуальных веществ. Большинство из них флюоресцируют в УФ-свете и являются фенольными соединениями. Неустойчивый сорт Карабурну отличается от Рипариа х Рупестрис 101-14 большим набором флавоноидных соединений, которые располагаются в центре хроматограммы. В листьях обоих сортов в наибольшем количестве содержится вещество 1, идентифицированное как флавонол-3-моногликозид-изокверцитрин; 2 — галлокатехин; 3 — флавоногликозид; 22 — кверцетин (рис. 3).
Различия в качественном составе всего дубильного комплекса листьев изученных сортов четко наблюдаются также при сравнении количества различных фракций дубильных веществ (Гадиев, Зотов, 1971а).
Содержание дубильных веществ в здоровых и пораженных мильдью листьях винограда (Mg/g сухих листьев)

Таблица 11
Таблица 10
Влияние ингибиторов роста на развитие спор Botrytis cinerea
| Вещества | Пораженные ткани листьев | Кора побегов (ноябрь) | ||
длина ростовых трубок, μ x±m | % | длина ростовых трубок, μ х±m | % | |
Контроль (вода) | 1,2+-2,7 | 100 | 25,7±4,1 | 100 |
Вещество № 4 | 9,4 ±2,6 | 77 | 15,3 ±3,3 | 56 |
5 | 4,5 ±1,2 | 37 | 5,7 ±0,67 | 22 |
6 | 8,0±0,01 | 6,7 | 2,5 ±0,37 | 9 |
6а | 5,0 ±1,4 | 42 | — |
|
6б | 6,2±0,2 | 52 | 10,8±1,6 | 42 |
Комплекс фенолкарбоновых кислот | 14,4 ±4,7 | 120 | — |
|
Раствор ИУК 200 μg/cm3 | 17,6±2,0 | 147 | — |
|
Данные табл. 10 показывают, что устойчивый сорт винограда характеризуется повышенным содержанием легкоокисляемых простых водно-растворимых соединений. Под влиянием инфекции в тканях восприимчивого сорта общее количество дубильного комплекса снижается, а у устойчивого сорта, напротив, происходит увеличение содержания всех фракций дубильных веществ.
Танинам придается определенное значение в устойчивости растений. Растворы танинов инактивируют ряд ферментов (Опарин, Купленская , 1935). Круикчанк и Перрин (1968) считают, что токсическое действие танинов обусловливается ферментативными превращениями и, кроме того, зависит от типа структуры самих танинов.
Таким образом, устойчивый сорт Рипариа х Рупестрис 101-14 характеризуется наиболее высоким уровнем активации пероксидазы и интенсивности дыхания, в результате чего в очаге инфекции в большом количестве образуются вещества липидной и фенольной природы, обладающие токсическими свойствами.
Сходным же образом реагирует виноградное растение в ответ на воздействие другого облигатного паразита — оидиума (Uncinula necator) и факультативного паразита Botrytis cinerea (Гадиев, 1969; Зотов, Плискановский, 1973).
Вышеописанные антибиотические вещества инфицированных тканей винограда нельзя отнести к фитоалексинам, так как подобные же соединения обнаружены в коре и почках виноградной лозы в период покоя. Однако это не отрицает возможности образования специфических продуктов взаимодействия виноградного растения и возбудителя мильдью — фитоалексинов, и наличия их среди вышеуказанных защитных веществ, выделение которых требует детального и углубленного изучения.
Последующие исследования показали, что некротизированные ткани листьев и покоящиеся почки, кора (луб) содержат сходные соединения, обладающие сине-голубым свечением в УФ-свете, близкими значениями Rf (0,01—0,4; 15%-ная уксусная кислота) и одинаковыми полосами поглощения в области 280—295 nm (Гадиев, 1975). Вещества № 5, 6, 6а, 6б (рис. 4) из покоящейся коры Рипариа х Рупестрис 101-14 (декабрь) обладают свойствами таковых из некротических тканей листьев. Дают положительные реакции с ДСК (№ 6, 6а) и родамином G (№ 5, 6б). Водные растворы веществ № 4, 5, 6, 6а, 6б вызывают торможение роста спор В. cinerea. В табл. 11 представлено действие ингибиторов роста из некротических тканей листьев и коры побегов Ри пария х Рупестрис 101-14 на рост спор В. cinerea. Все эти соединения оказывают тормозящее действие на рост спор. Наибольшим тормозящим эффектом обладают вещества 5, 6, 6а.
Максимальное содержание ингибиторов роста в коре и почках виноградной лозы наблюдается в период глубокого покоя (ноябрь — декабрь), а в период вегетации они не обнаруживаются. Содержание этих соединений в период покоя коррелирует с морозоустойчивостью виноградной лозы. Наибольшее количество ингибиторов роста содержится у американских диких видов (подвоев), американо-европейских и амурско-европейских гибридов. Наименьшим содержанием этих соединений в период покоя отличаются такие европейские сорта, как Карабурну, Чауш, Альфонс Лавалле, Корна нягра и др.
В отличие от вегетирующих зеленых частей и некротических тканей листьев, которые содержат большой набор флавоноловых соединений, виноградная лоза в период покоя (луб) содержит значительное количество катехинов.
Ингибирующее действие фенольных и липидных соединений, образующихся в инфицированных тканях винограда, а также при наступлении покоя, несомненно, является одним из важных защитных механизмов виноградного растения против повреждающих факторов различной природы. Эти соединения следует отнести к Эндогенным регуляторам роста. Общеизвестно, что фенольные соединения несут в растениях функции регуляторов роста (Кефели, Турецкая, 1965; Кефели, 1970, 1974; Wareing, 1965; Верзилов, Плотникова, 1972; Метлицкий с сотр., 1969; Пашкарь, 1970).
Важную роль в устойчивости растений к экстремальным факторам выполняют также липиды, являясь составными компонентами клеточных структур и мембран. Показано, что пониженные температуры вызывают в тканях винограда интенсивный синтез липопротеидов, значительная часть которых состоит из фосфолипидов.
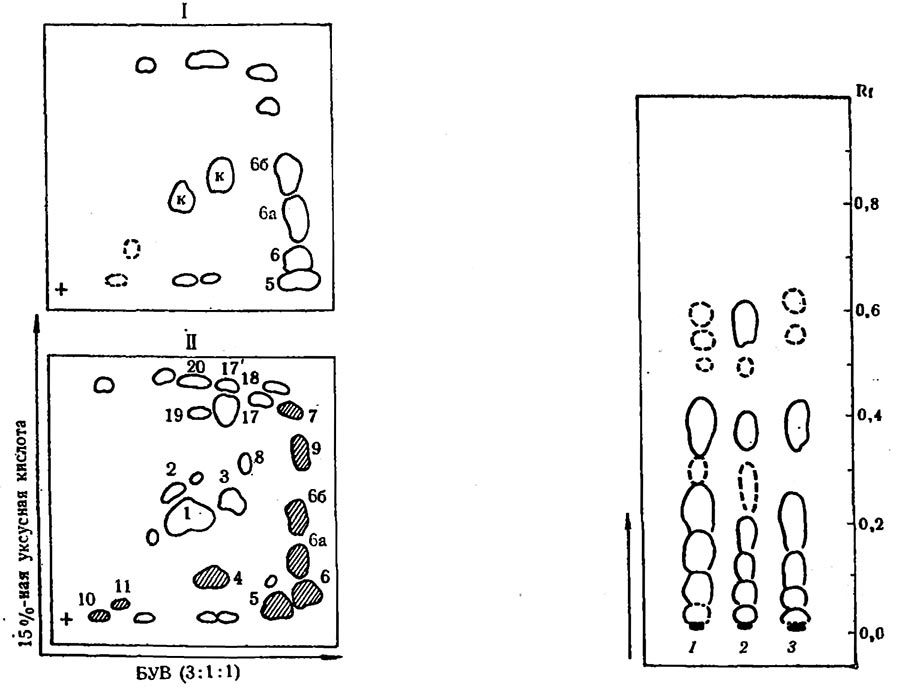
Рис. 4. Схема сравнительной хроматографии этанольных экстрактов различных органов устойчивого сорта Рипариа х Рупестрис 101-14
I — кора (луб) побегов за декабрь; II — поражение мильдью ткани листьев (некрозы); компоненты 4, 5, 6,6а, 6б, 7, 9, 10, 11 — вещества липидной и фенольной природы; 1, 2, 3, к — флавоноиды и катехины
Рис. 5. Схема одномерной хроматографии этанольных экстрактов покоящихся почек винограда (декабрь)
1— Рипариа X Рупестрис 101-14; 2 — Карабурну; 3 — Сейбель 1000. Растворитель — 15%-ная уксусная кислота
Они участвуют в переносе электронов, регулируют проницаемость мембран и поддерживают окислительное фосфорилирование которое приобретает решающее значение в зимний период (Дограмаджян, Аветисян, 1975). Значительное повышение фракции фосфолипидов у виноградной лозы в зимний период обеспечивает сохранность клеточных структур у устойчивых сортов при действии низких температур (Марутян, 1974). Согласно С. Е. Бреслеру, ферментативная активность мембранных белков в экстремальных условиях зависит от гидрофобных взаимодействий белка с фосфолипидами, при этом фосфолипиды выполняют функцию регуляторов конформационной устойчивости белка (цит. по Родионову с сотр., 1971).
Описанные выше ингибиторы роста фенольной и липидной природы являются общими ингибиторами роста, которые обусловливают комплексную устойчивость к грибным болезням и низким температурам.
Это подтверждает существующее мнение о менее выраженной специфичности иммунитета у растений, чем у животных.
На большом количестве сортов (более 100) винограда различного происхождения при помощи одномерной хроматографии на бумаге проверена зависимость между уровнем содержания эндогенных ингибиторов роста и степенью устойчивости. Хроматография на бумаге позволяет улавливать тонкие изменения в содержании этих ингибиторов роста (рис. 5). Эта закономерность принята в основу для разработки косвенного метода определения мильдью устойчивости винограда по относительному содержанию ингибиторов роста в покоящихся почках (Гадиев с сотр., 1976). Оценку степени устойчивости проводят по 5-балльной системе. Критерием служат размер пятен ингибиторов (Rf 0,01—0,45, 15 %-ная уксусная кислота) и интенсивность их свечения, которые находятся в прямой зависимости от степени устойчивости, а также наличие пятен наиболее подвижных фенол- карбоновых кислот (Rf 0,6—0,8), находящихся в обратной зависимости.
Баллом 1 оценивают устойчивый подвойный сорт Рипариа х Рупестрис 101-14, хроматограммы которого характеризуются наличием больших пятен ингибиторов роста, интенсивным свечением их в УФ-свете и незначительным содержанием фенолкарбоновых кислот. Эталоном сорта, устойчивость которого оценивается в 5 баллов, служит наиболее восприимчивый сорт Карабурну, хроматограммы которого отличаются слабосветящимися небольших размеров пятнами ингибиторов роста и содержанием значительного количества фенолкарбоновых кислот. Баллами 2, 3, 4 оценивают сорта, отличающиеся по степени устойчивости от двух крайних сортов и занимающие промежуточное положение.
Достоверность метода проверена на предварительно выделенных сеянцах гибридной комбинации Датье де Сан Валье х Декоративный. Проведена параллельная оценка степени мильдью устойчивости растений, выращенных из черенков этих же гибридных сеянцев в условиях жесткого инфекционного фона, по типу поражения листьев и интенсивности спороношения гриба. Сопряженность этих двух оценок выражается коэффициентом корреляции r=0,78+0,07.
Изученные сеянцы характеризуются большим разнообразием по степени мильдью устойчивости и оценены баллами от 3 до 5. Хроматографический метод позволяет выявлять незначительные различия в качественном составе метаболитов покоящихся почек сеянцев в пределах одной гибридной комбинации и может служить достоверным косвенным методом оценки мильдью- устойчивости винограда в селекционной работе.
Поскольку ингибиторы роста являются неспецифическими защитными веществами виноградного растения при действии различных повреждающих факторов (мильдью, оидиум, серая гниль, мороз), то эти соединения, по-видимому, представляют собой один из биохимических показателей комплексной устойчивости винограда к указанным факторам.
В настоящее время исследованиями ряда авторов (Вердеревский, Войтович, 1974; Гузун, Журавель, 1974; Айвазян, 1974) показана реальная возможность получения сортов винограда, обладающих комплексной устойчивостью к морозу, грибным болезням и филлоксере. При этом имеет место независимое наследование признаков морозо- и мильдью устойчивости и высокого качества ягод.
Для ускорения селекционной работы и повышения ее результативности необходимо усилить исследования молекулярно-генетических основ иммунитета, совершенствовать методы оценки устойчивости гибридных сеянцев.