Вопрос о каталитических системах и химических соединениях, синтезируемых патогенными микроорганизмами и обуславливающих их повреждающее действие на растение, включая гибель последнего, изучается на протяжении многих десятков лет.
Накопленные в этих исследованиях материалы свидетельствуют о большой зависимости свойств патогенов от типа их питания(сапрофиты, полупаразиты, облигатные паразиты и т. п.), а также от состава питательной среды. В число этих факторов должны быть включены и свойства растения, на котором они паразитируют. Этот вопрос более подробно освещается ниже. Огромное значение имеет ярко выраженная у патогенных микроорганизмов способность к образованию т. наз. адаптивных ферментов. Этим термином обозначаются ферменты, образование которых стимулируется теми или иными составными веществами питательной смеси. Специфика этих ферментов состоит в том, что они регулируют превращения тех химических соединений, под непосредственным воздействием которых они сами образовались. Механизмы этого стимулирующего влияния окончательно не выяснены. Согласно наиболее принятым в настоящее время взглядам соотношения между отдельными различными белками, включая и каталитически активные белки клетки (ферменты), регулируются двумя группами специфических агентов—репрессорами и дерепрессорами. Функции репрессоров заключаются в подавлении процессов биосинтеза белков, тогда как дерепрессоры снимают ингибирующее действие репрессоров. Под влиянием дерепрессоров в клетке либо стимулируется биосинтез раннее синтезировавшихся в незначительных количествах ферментов, либо создаются условия для синтеза ферментов, которых до этого в клетке совсем не было (синтез de novo). В роли репрессоров и дерепрессоров могут выступать многие, причем весьма разнообразные агенты химической и физической природы, включая компоненты питательной смеси, и метаболиты микроорганизмов, а также продукты обмена инфицированных клеток растения-хозяина.
В целом можно считать установленным, что возбудители заболеваний растений вооружены мощным ферментативным аппаратом, с участием компонентов которого могут осуществляться превращения практически всех содержащихся в тканях зеленого растения соединений. Это относится к различным группам гидролитических ферментов, включая и катализаторы, участвующие в разложении элементов покровных тканей растения (целлюлоза, гемицеллюлозы, пектиновые вещества, соединения группы лигнинов и их производных и др.). При участии этих ферментов преодолеваются механические барьеры на пути прорастающих спор, осуществляются первые этапы заражения — внедрение паразита внутрь клетки хозяина, установление непосредственного контакта и взаимодействия с содержащейся в ней протоплазмой.
Фитопатогенные микроорганизмы располагают различного рода протеолитическими ферментами, ферментами нуклеинового обмена, эстеразами, глюкозидазами, ферментами, расщепляющими полимерные формы углеводов и т. д. Весьма сложным является также состав ферментов окислительно-восстановительного комплекса. В нем широко представлены системы активации кислорода, дегидрогеназы, достаточно сложный комплекс коферментов пиридиновой и флавиновой природы, системы переносчиков, участвующих в транспорте электронов, и т. д. Фитопатогены располагают также полноценными системами, связанными с процессами окислительного фосфорилирования и синтеза АТФ. Следует подчеркнуть, вместе с тем, исключительно высоко выраженную у микроорганизмов динамичность отдельных ферментных систем и энзиматического аппарата в целом. Именно этим и обусловлена свойственная микроорганизмам пластичность и способность к адаптации, которыми определяется их способность к использованию в качестве энергетического и пластического материала самых разнообразных соединений.
К основным средствам воздействия патогена на ткани растения-хозяина относится сложный комплекс соединений, объединяемых термином "токсины". Как известно, в медицинской и ветеринарной практике токсинами называют индивидуальные соединения, специфические для определенных патогенных бактерий, которые и вызывают соответствующее заболевание организма. В фитопатологии этот термин относят обычно не к какому-либо определенному веществу, а к комплексу соединений, являющихся продуктами обмена возбудителя и выделяемых им при выращивании на синтетической питательной среде, а также при внедрении в живую клетку хозяина. Учитывая зависимость метаболизма патогена от состава питательной среды, нетрудно видеть, что состав "токсина" в том и другом случаях не может не быть различным. Некоторые авторы включают в понятие „токсин" также соединения, которые синтезируются клетками самого хозяина под влиянием внедрившегося в них патогена. Эту точку зрения последовательно развивает английский ученый Вуд, согласно которому токсином следует называть весь комплекс соединений инфицированной клетки, независимо от их происхождения. Обязательным их свойством должна служить способность нарушать нормальное функционирование протопласта, включая и гибель последнего. Вопрос о химической природе и механизме действия соединений, входящих в комплекс токсинов, весьма сложный. Можно считать установленным, что токсичным действием обладают соединения самой различной химической природы. К ним относятся разнообразные органические кислоты (щавелевая, лимонная, виннокаменная, фумаровая, а также гликолевая, глиоксилевая, молочная и др.), представляющие собой промежуточные продукты кислородного дыхания, либо конечные продукты брожения. Наряду с этими кислотами, действие которых, как правило, неспецифично, фитопатогенные грибы синтезируют и ряд кислот, обладающих весьма высокой и притом специфической физиологической активностью. Примером может служить альтерналиевая кислота, подавляющая прорастание семян и растений семейства пасленовых, фомовая кислота, выделенная из культурального фильтра гриба Phomamedicaginis, подавляющая прорастание семян льна, люцерны, рапса, кресс-салата и др. В основе физиологического действия этой кислоты лежит, главным образом, подавление функций ряда ферментов из группы дегидрогеназ.
Весьма активным токсическим действием обладают некоторые белковые соединения (например, альбумины), и, в особенности, продукты их распада (полипептиды, аминокислоты, аммиак, амиды, триметиламин и др.). Соединения этой группы ответственны большей частью за образование в инфицированных тканях некротических пятен.
Токсическое воздействие на ткани растения оказывают также некоторые из синтезируемых патогенами полисахариды. Они подавляют рост и синтез хлорофилла, вызывают увядание тканей, образование некротических пятен и т. п.).
Как правило, токсическое действие перечисленных выше соединений не отличается строгой специфичностью, а зачастую и вовсе неспецифично. Наряду с этим, известен ряд соединений, токсическое действие которых в той или иной мере специфично. Так, к числу высокотоксичных соединений может быть отнесена фузариновая кислота и обычно сопутствующие ей дипентид-ликомаразмин, а также вазинфуксарин.
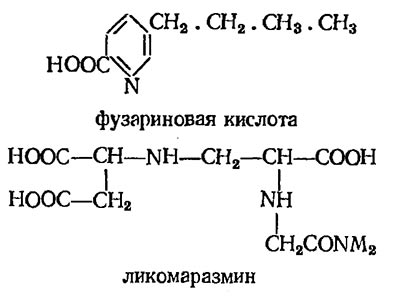
Биосинтез этих соединений характерен для грибов рода Fusarium — возбудителей болезни увядания большого числа различных растений. Доказано, что в основе их токсического действия лежит ингибирование функций важнейшей группы ферментов, связанных с энергообменом клетки (главным образом порфирин-производных). Отсюда следует ряд вторичных эффектов (задержка роста, в первую очередь, корней, образование некрозов на кончиках листьев, резкие нарушения водообмена, формирования и функционирования фотосинтетического аппарата и др.) Сильным токсическим действием обладают и некоторые полипептиды. Так же, как и в случае дипептидов, их токсичность обусловлена действием на системы дыхания, ультраструктуру протопласта и органоидов, на молекулярную организацию мембранных образований и обусловленную ею проницаемость последних и т. д.
Уже из этого краткого обзора следует, что фитопатогенные микроорганизмы обладают обширным арсеналом средств воздействия на растения- хозяина, которое направлено на нарушение нормального для его тканей течения процессов обмена веществ.
Естественно, что характер оказываемого микроорганизмами воздействия в большей мере зависит от их общей физиологической активности, которой и определяется вирулентность, патогенность различных видов возбудителей, а также их рас, штаммов и т. д. Исследования причин различной патогенности этих представителей проводится в разных направлениях. Одно из них состоит в изучении особенностей окислительного аппарата. Приведем в качестве примера материалы, полученные при сравнительном изучении вирулентного (№ 15) и авирулентного (№ 61) штаммов гриба Fusarium oxysporum vasinfeetum, возбудителя фузариазного вилта хлопчатника. Не останавливаясь на деталях, отметим лишь, что в структуре окислительного аппарата выявились четкие различия, тесно связанные с патогенностью штаммов. Высокая активность металлопротеидов и ферментов флавиновой природы и незначительная доля гексозомонофосфатного цикла — таковы обнаруженные сотрудниками кафедры физиологии растений МГУ Е. Р. Карташевой и Н. В. Гужовой основные особенности окислительной системы, которыми, по-видимому, и обусловлена большая патогенность гриба штамма № 15. Незначительная доля неактивных форм производных рибофлавина и, наконец, высокая активность ферментов пентозофосфатного цикла характеризуют авирулентный штамм.
С этими данными хорошо согласуются результаты многолетних работ советского исследователя З. Э. Бэккер, которая показала, что механизм токсического действия основного активного метаболита гриба Fusarium oxysporum vasifectum — фузариновой кислоты состоит в блокировании действия коферментов — производных никотиновой кислоты (НАД, НАДФ). Особенно высокой токсичностью обладает 3-д-бутилпиридин, образующийся из фузариновой кислоты путем ее декарбоксилирования. Из этих данных следует, что высокое содержание никотиновой кислоты может служить фактором, устраняющим или во всяком случае ослабляющим токсичность фузариновой кислоты. Тем самым подчеркивается связь между вирулентностью гриба Fusarium и его энзиматической системой, в первую очередь, ферментами, молекулы которых строятся при участии производных витаминов группы В.
Для выяснения возможных причин различий между этими двумя штаммами грибов в интенсивности порфиринового обмена были исследованы особенности реакции грибов на условия снабжения железом, которое, как известно, является одним из регуляторов биосинтеза порфириновых соединений. Так, от наличия железа в питательной среде зависит образование аминолевулиновой кислоты — промежуточного продукта на пути к синтезу пирролов. От снабжения железом зависит также превращение копрогена в протопорфирин. Установлено, что содержание в мицелии грибов протогема, активность каталазы и цитохромоксидазы у вирулентного штамма мало меняются в зависимости от уровня содержания железа в среде, тогда как у авирулентного штамма активность порфиринсодержащих ферментов и содержание гема при увеличении содержания Fe в среде возрастали в два и более раза. При увеличении дозы железа в десять раз содержание протогема и активность порфиринсодержащих ферментов у авирулентного штамма гриба весьма значительно возрастали.
Интересные данные получены при изучении АТФ-азной активности митохондриальных мембран у этих штаммов гриба. В отличие от обычной АТФ-азы, осуществляющей расщепление АТФ, АТФ-аза мембран митохондрий обладает функциями фермента, активирующего процессы биосинтеза АТФ, т. е. выполняет роль АТФ-синтетазы. Н. В. Гужова установила, что у вирулентного штамма гриба активность этого фермента почти в два раза выше, чем у авирулентного (соответственно 7,8 и 4,7). Эта закономерность подтвердилась при изучении функциональной активности митохондрий, изолированных из мицелия обоих штаммов гриба. У вирулентного штамма эффективность процессов окислительного фосфорилирования оказалась на 30—40 % выше, чем у авирулентного. Значительно более высоким является у этого штамма и содержание АТФ в мицелии.
Выяснилось, что по общему содержанию адениловых и никотинамидных: коферментов в клетке вирулентный штамм возбудителя вертициллезного вилта хлопчатника превосходит авирулентный. В особенности отчетливо характеризует это преимущество величина общего энергетического заряда изолированных митохондрий.
Таблица 1
Некоторые показатели энергосистем вирулентного и авирулентного штаммов гриба

Весьма показательна также величина отношения восстановленных форм нуклеотидов к окисленным, характеризующая уровень восстановленности этой системы. У вирулентного штамма это отношение в 1,5—2,5 раза выше, чем у авирулентного. Для вирулентного штамма Verticillium характерна, кроме того, более высокая по сравнению с вирулентным активность глюкозо-6-фосфат-дегидрогеназы — фермента, занимающего ключевое место в пентозо-фосфатном цикле дыхания (табл. 1).
Высокая энергетическая эффективность окислительных процессов обусловлена особенностями митохондриального аппарата.
Совокупность рассмотренных данных убеждает в том, что такие показатели энергообмена, как общая структура окислительного аппарата, состав ферментов и их изозимные спектры, природа переносчиков электронов, соотношение путей окисления и его энергетическая эффективность (коэффициенты P/О, АДФ/О), могут служить достаточно надежным ориентиром для суждения об общей физиологической активности возбудителя, его вирулентности. Эти показатели могут быть с успехом использованы в селекционной работе, при выведении сортов хлопчатника, устойчивых к патогенным агрессивным штаммам возбудителей вилта.
Весьма перспективными для целей диагностики уровня вирулентности возбудителей заболевания, равно как и устойчивости растения-хозяина к действию инфекции оказались показатели, регистрируемые методами биохемолюминесценции. Сюда относятся явления сверхслабого свечения (ССС), а также длительного послесвечения.
Сверхслабое свечение представляет собой спонтанное световое излучение живых клеток в видимой области спектра. Интенсивность этого свечения настолько низкая, что она не может быть оценена невооруженным глазом. В связи с этим для регистрации ССС используется специальная аппаратура из группы т. наз. фотоумножителей. ССС связано с окислением некоторых соединений клетки, главным образом липидов и фенолов. Оно наблюдается только при обязательном присутствии кислорода. Установлено, что в механизме ССС участвуют активные формы кислорода (свободные радикалы кислорода, возбужденный кислород, перекисные соединения и др.).
Обнаружено, что повреждающее воздействие на живую клетку факторов самой различной природы вызывает "вспышку" — усиление ССС различной продолжительности (от нескольких секунд до нескольких часов). Вспышка сопровождается увеличением концентрации свободных радикалов и накоплением продуктов окисления липидов и фенолов.
Причины вспышки, возникающей под воздействием различных факторов, различны. Однако в целом можно сказать, что этот эффект обусловлен нарушением состояния физиологических систем, защищающих клетку от побочного токсического действия кислорода. В результате происходят окисление мембранных липидов и другие патологические процессы, часть которых сопровождается свечением.
Установлено, что характер изменения ССС растений зависит от степени их устойчивости к определенному повреждающему воздействию (высокие и низкие температуры, обезвоживание и др.). Поэтому хемилюминесцентный метод уже эффективно используется для оценки устойчивости сельскохозяйственных культур к засухе, морозу и другим неблагоприятным воздействиям.
Следует учитывать, что метод ССС применим лишь в работах, объектом которых служат ткани, не содержащие зеленых и желтых пигментов. В исследованиях же с пигментированными тканями, в том числе с листьями, более применим метод, получивший название длительного послесвечения (ДСП). Сущность его состоит в следующем. Если лист осветить видимым светом, а затем поместить в темную камеру, то при помощи фотоэлектронного умножителя можно после прекращения освещения обнаружить затухающее в течение нескольких минут свечение в красной области спектра. Это явление обнаружено Стреллером и Арнольдом в 1951 г. и получило название ДПС.
Исследования показали, что послесвечение является универсальным свойством фотосинтезирующих организмов. Оно возникает в результате частичного обращения первичных этапов запасания энергии поглощаемых пигментами листа квантов света. В результате взаимодействия первично окисленного и первично восстановленного продуктов при фотосинтезе может возникнуть возбужденное состояние хлорофилла, дезактивация которого приводит к выделению квантов флюоресценции. Последняя и обнаруживается в виде послесвечения. Следует считать установленным также, что послесвечение — не побочное явление. Его параметры тесно связаны с ходом фотосинтеза и, прежде всего, с процессом фотофосфорилирования.
ДПС регистрируется на специальных установках, позволяющих чередовать импульсы возбуждающего света с периодами темноты, во время которых и наблюдается послесвечение. Исследования особенностей ДПС и его основных параметров позволили выявить вполне четкие различия в характере реагирования на заражение устойчивых и неустойчивых к вилту форм и сортов хлопчатника.
Установлено, что у неустойчивых сортов уже на первых этапах заболевания отмечается нарушение процессов фотофосфорилирования. Белковые компоненты хлоропластов у этих форм растений значительно менее подвижны, менее способны к конформационным изменениям своей структуры под действием света, что, в свою очередь, резко снижает эффективность функций Хлоропластов.
У устойчивых сортов нарушения фотофосфорилирования здесь не имеют места. Эти закономерности подтвердились при анализе значительного числа сортов различной устойчивости, что позволяет рассматривать ДПС как один из показателей, который может быть использован в селекционной работе.