Рассмотренные выше данные подчеркивают, что вызываемое патогенными микроорганизмами воздействие непосредственно затрагивает всю совокупность глубинных, интимных механизмов, регулирующих обмен здоровой ткани растения. Эти агенты вызывают различные нарушения функций растения-хозяина, дезорганизуют, извращают его нормальный метаболизм. Интенсивность и направленность этих нарушений обусловлены генетически. Они зависят от устойчивости хозяина, вирулентности патогена и носят в целом адаптивный характер.
Вместе с тем ход процессов патогенеза исключительно тесно связан с условиями, в которых протекает взаимодействие партнеров. При анализе этих процессов нельзя упускать из виду то, что их ход определяется исторически сложившейся взаимной приспособленностью растения и паразита. Отсюда следует, что характер процессов патогенеза должен быть обусловлен не односторонним влиянием возбудителя на ткани хозяина, а взаимовлиянием этих организмов. Иными словами, взаимоотношения последних осуществляются в полном соответствии с принципом прямой и обратной связи.
О правильности этого принципиально важного положения свидетельствуют обширные экспериментальные данные, полученные на кафедре физиологии растений МГУ при детальном изучении влияния, оказываемого растением-хозяином на регуляторные системы возбудителя.
Так, Е. Р. Карташова установила, что мицелий одного из возбудителей вилта хлопчатника гриба фузариума, выращенного на синтетической питательной среде и затем перенесенного на 6 h на вытяжку из тканей хлопчатника, по ряду весьма важных показателей отличается от выращенного только на искусственной питательной среде. Характер реагирования на состав среды был у штаммов различной вирулентности неодинаков (рис. 5). У вирулентного штамма гриба (№ 15) инкубация на растительной вытяжке сопровождается резкой активацией его общей физиологической активности: включение меченого углерода глюкозы в биомассу гриба, повышение общего уровня окислительных процессов и их энергетической эффективности и, что особенно важно, образование новых компонентов в изозимном спектре малатдегидрогеназы.
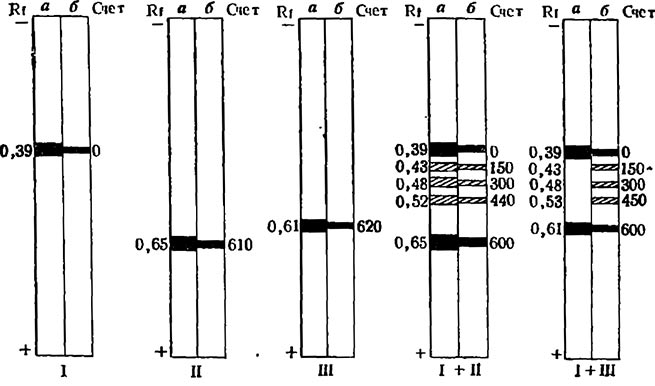
Рис. 5. Электрофореграммы грибных изозимов малатдегидрогеназы, полученных путем совместной диссоциации и реассоцяа- ции исходных изозимов фермента хлопчатника и гриба
исходные изозимы растения (I), 15-го штамма гриба (II) и 61-го штамма гриба (Ш);гибридизированные смеси Ι+Π и I-f-III;a—окраска геля на активность МДГ; б — окраска геля на белок; счет — радиоактивность фракции, рассчитанная в imp/min на 100 mg белка
Противоположна картина у авирулентного штамма (№ 61): включение 14С уменьшалось, новые изозимы малатдегидрогеназы (МДГ) не образовывались. Такого же рода данные накоплены Е. В. Юриной при изучении изозимных спектров пероксидазы уредоспор одного и того же вида ржавчины (Puccinia graminis), полученных от грибов, пассированных на различных сортах пшеницы. К аналогичным результатам пришли в опытах, где сопоставляли экзимограммы пероксидазы уредоспор ржавчины, выращенной на экстрактах пшеницы и ячменя.
С помощью препаративного электрофореза в полиакриламидном геле были изолированы индивидуальные изозимы из препаратов МДГ, выделенных из корней хлопчатника и мицелия вирулентного (№ 15) и авирулентного (№ 61) штаммов гриба Fusarium oxysporium vasinfectum. Предварительно препараты фермента были очищены фракционированием (NH4)2SO2 и хорматографией на сефадексе G-200. Диссоциация изозимных белков на субъединицы и реассоциация последних достигалась путем замораживания в 1 М растворе NaCl и последующего оттаивания. При электрофорезе диссоциированных и реассоциированных смесей изозимов МДГ вирулентного штамма и растения наблюдалось образование новых форм белка.
Для исследования структуры новых белков фракций в исходные изозимы МДГ гриба вводился изотоп 14С. На электрофореграммах гибридизированных смесей радиоактивность распределялась между исходной полосой МДГ гриба и новыми белковыми фракциями. Иммунохимические исследования показали, что в структуре новых форм белка наряду с субъединицами МДГ гриба имеются структурные элементы изозима МДГ растения. Иными словами, образовывались гибридные формы белка (рис. 6).
Появление у новых молекулярных форм белка полос преципитации с сыворотками к изозимам МДГ гриба и хлопчатника свидетельствует о наличии в структуре этих белков антигенных детерминант, свойственных обеим исходным формам МДГ.
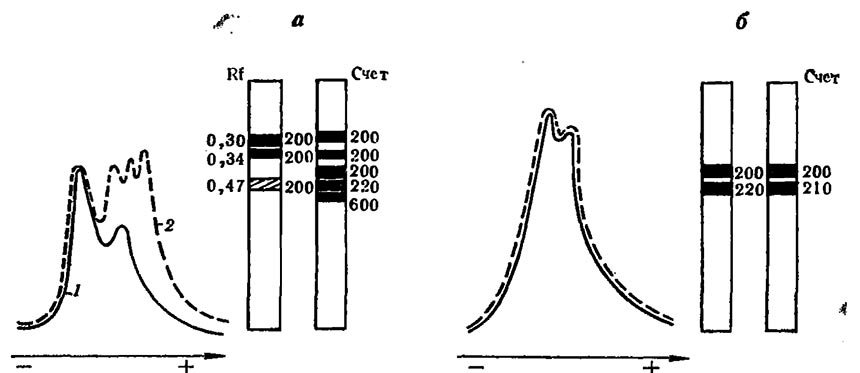
Рис. 6. Иэозимный спектр малатдегидрогеназы вирулентного (а) и авирулентного (б) штаммов гриба Fusarium oxysporum sp. vasinfectum под влиянием среды выращивания
1 — инкубация на искусственной питательной среде; 2—инкубация на вытяжке из корней; счет— радиоактивность полос МДГ, рассчитанная в imp/min на 100 mg белка
Это же следует из данных анализа аминокислотного состава гибридных полос фермента. Было установлено, что содержание остатков аминокислот в гибридных фракциях равнялось среднему от содержания их в исходных фракциях. Это опять-таки было справедливо как к гибридным полосам, обладавшим ферментативной активностью, так и к неактивным. Отсюда видно, что комплементарность субъединиц, позволяющая сохранить ферментативную активность, обусловлена не составом аминокислот, а в первую очередь последовательностью расположения последних в белковой молекуле и особенностями конформации ее вторичной и третичной структуры. Образовавшиеся de novo формы изозимов обладали ферментативной активностью только при условии, что при этом участвовали субъединицы МДГ вирулентного штамма. В случае авирулентного штамма эти гибридные формы белка ферментативной активностью не обладали. Следует особо подчеркнуть, что новых белковых полос не было, когда диссоциация и реассоциация изозимов МДГ гриба и хозяина проводились раздельно и смешивались только непосредственно перед электрофорезом.
Сопоставление физико-химических свойств изозимов одного и того же фермента хозяина и паразита позволяет получить представление о биологической совместимости этих организмов.
Дальнейшие исследования этой сложной проблемы представляют не только теоретический интерес, но открывают хорошие перспективы с точки зрения их использования в селекционной практике. В частности, их результаты должны найти применение при формировании комплекса штаммов возбудителей вилта хлопчатника на т. наз. провокационных фонах. Последние, как известно, широко используются в работах по созданию вилтоустойчивых сортов хлопчатника.
Из рассмотренных данных следует, что генеральная задача, стоящая перед сельскохозяйственной наукой и практикой в области иммунитета, может быть кратко сформулирована следующим образом: отыскание путей преодоления биологической совместимости растения-хозяина и возбудителя его заболевания.
В настоящее время основное внимание при решении этой задачи сосредоточено на создании форм растений с наследственно, генетически закрепленными свойствами иммунитета. На этом пути достигнуты и достигаются огромной возможности результаты. В особенности ценные материалы получены при вовлечении в селекцию диких, некультурных сородичей культурных растений, которые служат ценным источником генов, контролирующих иммунитет, к которым не адаптированы гены соответствующих агентов.
Что касается возбудителей, то они преодолевают устойчивость высшего растения путем возникновения в их наследственном аппарате генов вирулентности, совместимых с каждым геном устойчивости хозяина.
Как долго новый сорт сохраняет устойчивость, зависит от числа введенных в него генов, некомплементарных к генам вирулентности соответствующего возбудителя, а также от быстроты приобретения патогеном необходимых новых генов вирулентности.
Вся история селекции на иммунитет полна примеров того, как выведенный иммунный сорт через некоторое время становится поражаемым. В подавляющем большинстве случаев в основе этого явления лежит формирование новых биологических форм возбудителя (физиологические расы, штаммы и др.), преодолевших барьер несовместимости с созданными селекционерами иммунными формами растений.
Один из примеров — обстоятельная работа Штаббса. Изучая состав популяции желтой ржавчины в Голландии за последние двадцать лет, Штаббс обнаружил значительные изменения патогенности возбудителя, обусловленные накоплением соответствующих генов. Непосредственной причиной этих изменений является создание коллекционерами новых, устойчивых к прежним расам возбудителя сортов пшеницы.
Селекционер не может рассчитывать на то, что выведенный им иммунный сорт сохранит свои свойства как постоянный признак. Параллельно отбору искусственному в природе протекает отбор естественный, который закрепляет мельчайшие отклонения, оказавшиеся полезными для выживания паразита и тем самым способствующие возникновению новых рас с более совершенными свойствами. На это указывал еще Чарльз Дарвин.
Необходимо учитывать, кроме того, что темпы создания новых, устойчивых к тому или иному патогену сортов во много раз более медленны, чем скорость расообразовательных процессов у паразита.
Сорт—генетически обусловленная совокупность потенциальных свойств растения, реализация которых в большей мере зависит от конкретных условий, в которых оно развивается. Сорт — это интегрированная система, в которой компоненты генотипа проявляются лишь в определенных условиях среды. В связи с этим, наряду с имеющей непреходящее значение селекционной работой, в поле зрения растениеводов и селекционеров как весьма эффективное дополнение к ней должны быть включены мероприятия по воздействию на растения в онтогенезе, по повышению устойчивости в ходе индивидуальной жизни организма.
Данные, полученные при изучении интимных сторон взаимовлияния хозяина и паразита, высокая степень подвижности качественных и количественных параметров ферментативно активных белков клетки не оставляют сомнения в возможности такого рода воздействия.
Нарушение биологической совместимости партнеров в системе хозяин- паразит может быть достигнуто путем воздействия на энзиматический аппарат и обмен веществ в целом как хозяина, так и паразита.
Одним из эффективных путей такого воздействия, до настоящего времени недостаточно используемым, несомненно, может служить весь комплекс приемов агротехники, и, в первую очередь, условия минерального питания.
Имеющаяся по этому вопросу литература, в общем, очень немногочисленна, причем сведения, содержащиеся в ней, разноречивы, нередко противоречивы. Причина в том, что систематическому изучению, исходящему из современных теоретических представлений о биологической роли элементов минеральной пищи, в первую очередь, микроэлементов, эта проблема не подвергалась. Между тем, именно микроэлементы участвуют в формировании молекул тех групп ферментов, роль которых в регулировании процессов, протекающих в инфицированной клетке, выше подробно освещалась.
Всем указанным подчеркивается необходимость организации детального, а главное систематического изучения всего комплекса факторов, контролирующих процессы образования физиологических рас. Детальному изучению должна быть подвергнута вся сумма вопросов, касающихся природы механизмов, которыми обусловлена вирулентность, агрессивность, и, наконец, патогенные свойства возбудителей.
Исследования в этой области имеют не только большое теоретическое значение, но и весьма важны в практическом отношении. В успешном их развитии заинтересованы, прежде всего, селекционеры.
В настоящее время получило признание положение, согласно которому наиболее надежный путь борьбы с болезнями растений состоит в выведении устойчивых (иммунных) сортов. Одним из основных методов, используемых в этих целях, является испытание вновь создаваемых селекционером форм растений на т. наз. инфекционных (провокационных) фонах. Сущность этою приема заключается во внесении в почву определенных количеств инфекционного материала (культура гриба-возбудителя, выращенного в лабораторных условиях на той или иной среде, запахивание в почву больных растений и т. п.).
В особенности широко используется проверка на инфекционных фонах в селекции сельскохозяйственных культур на устойчивость к корневым болезням либо заболеваниям, осуществляемым микроорганизмами, проникающими в растение через корни. Все эти приемы страдают существенными недостатками, заключающимися в том, что они не позволяют контролировать количество вносимого в почву инфекционного материала и, что в особенности важно, качественного (расового) состава возбудителей.
Учитывая широкие пределы варьирования уровней физиологической активности (вирулентность, патогенность) возбудителей заболевания растений вилтом, различного типа корневыми гнилями и т. п., становится очевидной необходимость введения строгой объективной количественной и качественной оценки искусственных инфекционных полей. Именно этим целям и должны служить исследования, направленные на выяснение физиолого-биохимических основ патогенности, вирулентности возбудителей болезней. Их результаты должны стать фундаментом подбора форм патогенов при формировании инфекционных фонов.
Итак, патогенез представляет собой сложную совокупность физиолого- биохимических процессов. Их направленность, интенсивность и конечный эффект являются результатом взаимодействия управляющих механизмов антагонистичных биологических систем—ауто- и гетеротрофных организмов. Взаимное влияние этих организмов направлено на жизненно важные центры физиологической активности каждого из партнеров (системы биосинтеза белка, системы энергообмена).
При оценке значения систем энергообмена основное внимание должно быть уделено уровню общей эффективности их функционирования: процессам первичного использования солнечной энергии, ее преобразования, запасания и последующей реализации свободной энергии. Факторами, обеспечивающими высокую эффективность этих процессов, являются, в первую очередь, стабильность, устойчивость систем сопряжения, сочетающаяся с гибкостью этих систем, их пластичностью. Совокупностью этих свойств определяются уровень устойчивости растения, физиологическая активность возбудителя, а также характер взаимовлияния, их реагирования на обоюдное вмешательство.
Исследования на молекулярном уровне приносят новые доказательства правильности идеи Н. И. Вавилова о том, что свойства устойчивости зеленого растения, так же, как и свойства паразитизма, формировались при постоянном контролирующем взаимовлиянии этих организмов, что они эволюционировали сопряженно.
О ведущей роли белковых компонентов протопласта в формировании защитных свойств растения и вирулентности возбудителя свидетельствуют экспериментально установленные факты индуцированного образования в клетках партнеров иммунологически специфических белков, а также способность инфицированных клеток зеленого растения синтезировать белки, обладающие антивирусной активностью.
И в этом случае мы обязаны отдать дань глубокого уважения Н. И. Вавилову, который еще в 1940 г. в докладе на заседании Биоотделения АН СССР утверждал: "...при большем углублении в химическую природу белков плазмы, различающую виды и сорта, природа иммунитета станет более понятной".
Принципиальное значение приведенных выше фактов состоит в том, что они сближают представления о молекулярной природе защитных механизмов у растений с таковыми у животных, перебрасывают мост между ними. Нет сомнений в том, что дальнейшие исследования принесут новые доказательства правильности этой важной в общебиологическом отношении закономерности. Это будет находиться в полном соответствии с общепринятыми сейчас представлениями о близости, а во многих случаях — и полной идентичности принципов структурного и функционального построения всей живой материи на Земле.
С предельной четкостью эту идею сформулировал Джон Бернал в книге "Возникновение жизни". Он писал: "Все явления, изучаемые биологией, образуют непрерывную цепь событий, и каждое последующее звено нельзя объяснить, не принимая в расчет предыдущее. Единство жизни вытекает из всей ее истории и, следовательно, является отражением ее происхождения".