III.2. РОЛЬ СОЛНЕЧНОЙ РАДИАЦИИ В ФОРМИРОВАНИИ УРОЖАЯ ВИНОГРАДА
Оценивая роль солнечной радиации в продукционном процессе винограда, необходимо прежде всего остановиться на материалах, касающихся фотосинтеза — физиологического процесса, в результате которого из неорганических веществ и поглощенной листьями энергии ФАР создается органическая продукция.
Фотосинтез. Зависимость фотосинтеза от ФАР характеризуется так называемыми световыми кривыми фотосинтеза и газообмена, имеющими в общем виде гиперболическую форму. Ход световых кривых фотосинтеза характеризуется через так называемые «кардинальные точки» [249, 403]: компенсационный пункт (КП) — уровень ФАР, при котором интенсивность фотосинтеза равна интенсивности дыхания (поглощение CO2 уравновешивается его выделением); световое насыщение фотосинтеза—плотность потока ФАР, при которой достигается максимальная для данного сочетания внешних условий интенсивность фотосинтеза; уровень «плато» — состояние, когда при увеличении ФАР интенсивность фотосинтеза удерживается для данных условий на одном максимальном уровне; угол наклона световой кривой, который характеризует эффективность использования лучистой энергии на фотосинтез; радиация приспособления (ИРП) — плотность потока ФАР, при которой КПД газообмена достигает максимума [360].
Нa рис. 10, составленном нами по литературным источникам показаны световые кривые фотосинтеза листьев винограда. Данные эти получены в разных условиях и разными методами, потому трудно сопоставимы. Тем не менее они дают известное представление о закономерностях изменения интенсивности фотосинтеза листьев винограда в зависимости от освещенности. Пи верхней горизонтальной оси нанесены пересчитанные нами дополнительно энергетические эквиваленты люкса по ИР и ФАР.

Рис. 10. Световые кривые фотосинтеза винограда.
Световая кривая фотосинтеза листьев 1, полученная при температуре воздуха 27°С и относительной влажности воздуха 47% [44], показывает резкое увеличение фотосинтеза при увеличении освещенности до 12 тыс. лк со световым насыщением фотосинтеза при 40—50 тыс. лк и К.П в пределах 0,5—1,0 тыс. лк.
1 In световой кривой 2 при температуре воздуха 22—26° С и относительной влажности 65% [220] фотосинтез начинается около 2 тыс. лк и повышается по мере увеличения освещенности до (id тыс. лк. При влажности воздуха 50% световое насыщение поступало при 15—30 тыс. лк. На световой кривой фотосинтеза на открытом воздухе значениям KTI соответствует освещенность 1250 лк, а световое насыщение наступает в интервале 15-7-20ХЮ4 эрг/см2 (37—50 тыс. лк). В теплице К.П наступил при освещенности 500 лк, а полный свет оказывал ингибирующее действие на фотосинтез растений. Ассимиляционная способность зеленых ягод была значительно ниже, чем листьев (соответственно 50 мкл и 3,5 мл СO2/ч на 1 г сырого веса).
При кратковременных определениях [55] максимальная продуктивность фотосинтеза (2,7—3,0 г/(м2-сутки) наблюдалась на прямом свету при освещенности 33 тыс. лк. В зоне второго яруса листьев от основания побега (12—15 тыс. лк) продуктивность фотосинтеза составляла 1,0—1,6 г/(м2• сутки), а внутри кроны (2—4 тыс. лк)—всего 0,3—0,7 г/(м2• сутки). Штирбан и Тара [493] обнаружили максимум фотосинтеза листьев у сортов Фетяска, Мускат Оттонель и Траминер розовый при температуре 28—32° С и освещенности 37—47 тыс. лк. Бошьян (цит. по [418]) наблюдал максимум фотосинтеза при температуре воздуха 30° С и освещенности 60 тыс. лк.
По данным Погосяна и Мелконяна [285], в период цветения, роста ягод и созревания фотосинтез достигает максимума в 8—9 ч при 20—30 тыс. лк. В полуденные часы при освещенности 80—90 тыс. лк наблюдается депрессия фотосинтеза до нулевого уровня. В вечерние часы при освещенности 20—60 тыс. лк фотосинтез вновь повышается, хотя абсолютные его значения ниже утренних. В период полной зрелости ягод в связи с уменьшением поступления солнечной радиации кривая дневного хода фотосинтеза имеет одну вершину в полуденные часы при освещенности 20—40 тыс. лк.
Имеющиеся данные (табл. 8) показывают, что изменение освещенности резко сказывается на интенсивности накопления сухого вещества растением и величине ЧПФ [488].
Таблица 8
Влияние освещенности на накопление сухого вещества виноградным растением (по [488])
| Освещенность | Средняя освещенность в ясный день, тыс. лк | Прирост сухого вещества, растений | ЧПФ, | ||||
9 VII — 5 VIII | 6 VIII— 10 IX | 11 IX — 12 X | 9 VII— 5 VIII | 6 VIII — 10 IX | 11 IX | ||
Полная | 63 | 24,0 | 59,0 | 37,0 | 4,3 | 4,6 | 2,55 |
Частичное затенение | 19—63 | 14,4 | 28,0 | 19,0 | 2,4 | 2,3 | 1,85 |
Полное затенение | 2,6 | 3,2 | 6,2 | 5,6 | 0,6 | 0,5 | 0,51 |
Примечание. Сухой вес растения на 8 VII во всех вариантах равен 30 г.
По Шолису с сотр. [488], для максимального фотосинтеза необходима освещенность 20 тыс. лк.
Как показывают данные [450], из освещенных Солнцем листьев (40 тыс. лк) происходит значительный отток ассимиляции. Из находящихся внутри кроны или затененных листьев (НПО л к) отток ассимилятов практически отсутствует. В случае частичного попадания на листья внутри кроны солнечных лучей наблюдается перемещение ассимилятов из этих листьев и другие части куста. Нижний предел освещенности, при котором наблюдался отток ассимилятов, равнялся 1000 лк.
Видовые особенности реакции винограда на условия освещенности обнаружились Гейслером [433]. Компенсационный IIv11кт фотосинтеза изменялся от 300 лк у вида V. riparia до 1П00 лк у вида V. labrusca. Световое насыщение наступало соответственно при 10 и 28 тыс. лк. Сорта винограда V. vinifera занимали промежуточное положение.
Таким образом, данные показывают, что для винограда I манным образом V. vinifera) КП фотосинтеза соответствует уровню освещенности порядка 0,5—1,5 тыс. лк (25-75Х < 10-4 кал/(см2-мин) ФАР). Световое насыщение фотосинтеза винограда в зависимости от сорта и других факторов наступает и диапазоне освещенности 20—60 тыс. лк (0,1—0,3 кал/(см2Х »;. мин) ФАР); наибольшее число случаев приходится на освещенность порядка 30 тыс. лк (0,15 кал/(см2• мин) ФАР). Как известно, у большинства сельскохозяйственных растений КП фотосинтеза соответствует интенсивность ФАР, близкая к 0,01—0,025 кал/(см2• мин) (2—5 тыс. лк), а световое насыщение фотосинтеза наступает при интенсивности ФАР, близкой к 0,25—0,35 кал/(см2.мин) (50—70 тыс. лк) [149, 249].
Для винограда характерна большая пластичность, которая проявляется, в частности, в способности адаптации фотосинтеза к колебаниям температуры и освещенности. У сортов винограда Лидия, Черный сладкий и Амурский фотосинтез проходил без снижения и интервале 7—8 и 30—32° С при оптимуме 24—30° С [220]. У сорта Султана [452] полный свет оказывал ингибирующее действие на фотосинтез растений, адаптированных к условиям освещенности в теплице. При длительном пребывании на полном свету эффект ингибирования исчезал.
Высокая потенциальная способность фотосинтеза листьев винограда проявилась в опыте, где свет проникал через отверстие в кювете и освещал 8,5% площади листа. При освещенности 37 тыс. лк суммарный фотосинтез составил 25% полной площади листа. Если листья были обеспечены водой, они Могли выдержать около 3 ч температуру 48—49° С. При температуре выше 50° С наблюдалось интенсивное выделение CO2 и листья погибали. По Сэледжану [345], листья винограда, оставленные в течение 5 мин при температуре 50° С, сохранили способность к фотосинтезу. На большую способность адаптации винограда к температурным условиям указывает Илькун [136]. Фактические данные показывают, что средний для винограда оптимум температуры для фотосинтеза лежит в интервале 25—30° С; критической является температура 45° С [220, 447, 452, 493].
Оценивая приведенные данные по световым кривым фотосинтеза листьев винограда, а также значения кардинальных точек на этих кривых, следует отметить, что они могут приниматься как весьма приближенные, основанные на упрощенном подходе к определению функциональной зависимости фотосинтеза листьев от условий освещенности. Скорее здесь можно говорить не о световых кривых фотосинтеза виноградного растения, а о световых кривых тех листьев на растении, на которых проводились определения фотосинтеза (причем за короткий период). В отношении же целого растения определение световых кривых фотосинтеза листьев осложняется целым рядом факторов.
Прежде всего световые кривые фотосинтеза и газообмена листьев разных ярусов обычно неодинаковы. Например, в ценозах сплошного покрова листья, находящиеся в верхних хорошо освещенных ярусах, приспособлены к более сильному свету и обладают более высокой интенсивностью газообмена. В средних ярусах плато световых кривых несколько снижается, опускается и КП. Листья нижних ярусов приспособлены к более слабому свету [187, 216, 360, 406]. Отсюда можно ожидать, что неодинаковыми будут и световые кривые фотосинтеза листьев винограда, расположенных не только на разных ярусах, но и в разных частях кроны в пределах одного яруса: в наружной части кроны или внутри нее (на разной глубине).
Немаловажным является и то, что в процессе формирования ценоза разные листья оказываются в меняющихся условиях освещенности. По мере этих изменений фотосинтетический аппарат адаптируется к новым условиям [33, 80, 187, 268], а листьям каждого яруса соответствуют световые кривые, «настроенные» на максимальный КПД [360]. Иначе говоря, интенсивность фотосинтеза листьев может меняться непропорционально изменению освещенности; она может сохраняться на том же уровне при изменении (снижении) освещенности.
Как отмечает Тооминг [360], «предпосылкой создания фотосинтезирующих систем с высокой продуктивностью является установление формы зависимости кардинальных точек световых кривых газообмена от условий ФАР местообитания» (с. 148). Важным при этом является учет интенсивности радиации приспособления (ИРП) как кардинальной точки световой кривой, содержащей экологическую информацию о светолюбивости и экономичности дыхания данного вида растения.
Каждый вид растения имеет определенную возможность варьирования ИРП своих листьев в соответствии со средними условиями ФАР в ярусе произрастания. У сельскохозяйственных растений ИРП выше, чем у дикорастущих видов; у светолюбивых растений выше, чем у тенелюбивых. Сельскохозяйственные культуры с высокой продуктивностью (кукуруза, фасоль, ячмень, пшеница и др.) имеют значения ИРП, достигающие 0,05—0,1 кал/см2 для верхних ярусов.
Интерес представляет установление у винограда зависимости интенсивности фотосинтеза при световом насыщении и интенсивности дыхания от морфологических показателей листьев, таких, как удельная поверхностная плотность листьев (УПП), под которой подразумевается сухой вес единицы площади поверхности листа (г/дм2), и относительная площадь хлоропластов в листьях — суммарная площадь хлоропластов к расчете на единицу площади поверхности листа [360]. Эксперименты показывают, что генотипы и сорта с высокой максимальной интенсивностью фотосинтеза обладают и более высокой УПП листьев [401, 425, 426, 427].
Рост и плодоношение.
Роль солнечной радиации в продукционном процессе винограда не ограничивается фотосинтезом. От интенсивности света и продолжительности светлого и темного периода суток (фотопериода) зависят важные морфогенетические процессы, определяющие характер развития растений и его хозяйственную продуктивность. К их числу относятся, в частности, рост побегов, а также закладка и дифференциация генеративных органов в почках винограда.
Одним из первых исследований в этом направлении была работа Гакбарта и Шерца [434]. В пределах рода Vitis ими были отмечены следующие специфические для видов и сортов реакции: виды V. riparia, V. solonis и V. rupestris обнаруживали на коротком дне более сильное торможение роста, чем вид V. vinifera, т. е. европейские сорта оказались более устойчивыми к действию короткого дня, чем американские. Короткий день способствовал лучшему вызреванию побегов у всех видов рода Vitis. Длина междоузлий у разных видов на нормальном дне была всегда больше, чем на коротком, причем уменьшение длины междоузлий с уменьшением фотопериода у американских видов было выражено сильнее, чем у V. vinifera. У всех видов появление листьев (число узлов) подавлялось коротким днем в меньшей мере, чем прирост побегов. Вес корней увеличивался на коротком дне, несмотря на одновременное уменьшение длины побегов. Отношение между весом надземных и подземных органов у всех видов на коротком дне было примерно одинаковое; на нормальном дне оно изменялось и большей степени у американских видов.
Обнаружились отчетливо выраженные изменения формы листьев под влиянием разной освещенности. У американских видов длина и ширина листьев на коротком дне значительно уменьшались. У европейских сортов (Сильванер) размеры листа практически не менялись. В то же время на коротком дне и зависимости от сорта сужалась черешковая выемка, образовывались более широкие и тупые зубцы, усиливалось опушение, грубее становилась поверхность листа, укорачивались листовые черешки.
При ограниченном доступе солнечной радиации сильно подавляется рост губчатых, палисадных и эпидермальных клеток, что обусловливает образование более тонких пластинок листьев по сравнению с листьями кустов, выращиваемых на полном свету [386]. При адаптации винограда к условиям затенения образуются листья с более мезоморфной или мезоксероморфной структурой. По мере ослабления света увеличивается количество пластидных пигментов в листьях. Эта закономерность сильнее выражена в отношении хлорофилла, особенно хлорофилла «а», и слабее — в отношении каротиноидов [273,346,349].
По данным Аллевельдта [10. 410], с уменьшением фотопериода скорость роста побегов уменьшается у всех сортов винограда, причем влияние длины дня на прирост побегов и развитие листовой поверхности специфично у каждого сорта. Обнаруживается положительная корреляция между температурными условиями, продолжительностью солнечного сияния, интенсивностью образования цветков и скоростью их дифференциации в почках. Данные ряда авторов [93, 105, 291, 340] указывают на значительные изменения морфологических и физиологических признаков винограда на коротком дне, а также на значительные изменения в углеводном метаболизме надземных и подземных органов в сторону ускорения синтетических процессов. Большой интерес представляют данные о влиянии солнечной радиации на плодоносность почек винограда — формирование и дифференциацию в них соцветий.
Как отмечает Мозер [214], неблагоприятные условия погоды в мае и июне сказываются отрицательно на формировании урожая, так как в этот период у винограда в зимующих почках закладываются соцветия под урожай следующего года.
В условиях Австралии по 18-летним данным [412] обнаружилась тесная зависимость плодоносности почек от числа часов солнечного сияния в течение трех недель — с конца ноября до начала декабря, когда в почках происходит дифференциация зачатков соцветий. Для прогноза плодоносности почек получено уравнение регрессии, учитывающее число часов солнечного сияния и сумму дневных максимальных температур между 28 и 32° С для 20-дневного периода.
Результаты исследований с искусственным затенением глазков 1 показали, что солнечная радиация влияет на плодоносность почек путем непосредственного действия на глазок [61, 106, 448, 449, 467, 468] (рис. 11). Плодоносность почек повышается с увеличением общего количества поступившей радиации и не зависит от ее спектрального состава. Мей [467] связывает формирование зачаточных соцветий с развитием вегетативных компонентов глазка и подвергает сомнению положение о том, что фотопериодическое воздействие воспринимается листьями и передается от них к почкам, где соцветия закладываются.
1 У винограда «глазок» представляет собой сложное образование, состоящее из главной почки и нескольких замещающих почек.
Исследования выявили различную реакцию сортов на фотопериод [418, 419, 485] (рис. 12). По мнению Бутроза [419], несмотря на потребность в лучистой энергии, механизм плодоносности не идентичен механизму накопления сухого вещества растения. Если плодоносность почек зависит от общего количества часов с достаточно высокой облученностью, то накопление сухого вещества определяется главным образом напряженностью лучистой энергии и в меньшей степени — количеством часов ее воздействия.
Как известно, определение фактической плодоносности почек весной перед обрезкой кустов является одним из необходимых условий для установления оптимальной нагрузки кустов побегами и получения планируемого урожая винограда. Однако из-за трудоемкости существующих методов определения плодоносности почек (микро-скопирование глазков или их проращивание) работа эта не всегда может быть выполнена в нужном объеме и за короткий срок, особенно в зоне укрывного виноградарства и в районах, где глазки подвержены опасности повреждения поздними весенними заморозками. Обрезка же, проведенная без учета плодоносности побегов, часто приводит к значительному недобору урожая или же, напротив, может привести к «перегрузке» кустов гроздями и к ослаблению кустов.
Приведенные материалы показывают, что существует реальная возможность прогноза плодоносности почек винограда на основе коррелятивной связи между плодоносностью и количеством поступившей энергии солнечной радиации за определенный «критический период», когда в формирующихся глазках закладываются соцветия урожая следующего года. Разработка агроклиматических методов прогноза плодоносности винограда позволила бы в значительной степени упростить работу, связанную с оценкой состояния кустов, а следовательно, и упорядочить методы установления оптимальной нагрузки побегами для получения стабильных урожаев.
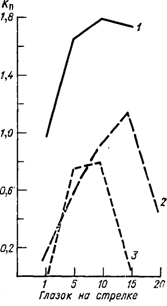
Рис. 11. Потенциальный коэффициент плодоносности глазка Кп сорта Болгар в зависимости от освещенности.
1 — полная освещенность, 2 — 01% от полной, 3 — 22% от полной (по [61]).
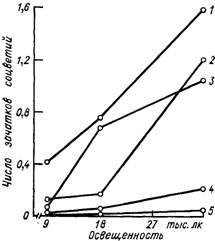
Рис. 12. Влияние интенсивности света на среднее число зачатков соцветий в почке (для базальных 12 почек на побеге). Продолжительность светового дня 16 ч при температуре 25° С.
Сорта: 1 — Рислинг рейнский, 2 — Шираз, 3 — Гордо, 4 — Оганез, 5 — Султана (по [419]).
В зависимости от условий перезимовки кустов и объема работы такие агроклиматические прогнозы можно было бы при необходимости корректировать данными о плодоносности, полученными существующими методами. Очевидно, что работа эта должна проводиться в разных природных зонах и охватывать как можно большее число сортов, имея в виду, что в дальнейшем они могут быть объединены в сортогруппы.
Качественный состав биопродуктов в ягодах.
Хозяйственно ценной частью виноградного растения являются ягоды, которые вместе с гребнем и ножкой гребня образуют гроздь. В ягодах винограда при полном созревании содержатся сахара, органические кислоты, минеральные и азотистые вещества, витамины и другие соединения. Окраска ягод красных, синих и черных сортов обусловлена пигментами-антоцианами, которые размещаются главным образом в клетках кожицы, а у некоторых сортов, кроме того, и в клеточном соке мякоти. На долю органических соединений приходится до 97% сухого вещества сока; минеральных веществ (зола) в ягодах 2—3% на сухое вещество.
Сахара в соке представлены в основном глюкозой и фруктозой; кроме моносахаров, в ягодах накапливается некоторое количество сахарозы. Кислоты виноградного сока представлены винной, яблочной, лимонной, аскорбиновой, фосфорной и в небольших количествах другими кислотами. На долю винной и яблочной приходится свыше 90% всех органических кислот [246]. Содержание сахаров и органических кислот в ягодах, а также соотношение между ними определяет вкусовые качества винограда и в значительной степени — направление использования его как продукции для виноделия.
Еще в ранних работах Любименко [179] была высказана мысль об использовании фактора «освещенности» для повышения урожая и изменения качества винограда. Выполненные им опыты с искусственным затенением [180] показали, что стимулирующее действие света на развитие ягод достигает наибольшей силы при некоторой вполне определенной напряженности освещения. При очень слабом доступе света, а также при избыточном свете развитие ягод задерживается. Наилучшее развитие ягод у сортов Шасла и Додреляби достигалось при некотором их затенении, причем сорт Шасла оказался более требовательным к свету, чем Додреляби. Благоприятное влияние слабого затенения сказывалось прежде всего на увеличении числа разнившихся ягод, хотя вес отдельных ягод из-за плотности грозди уменьшался. При оптимальных условиях затенения урожай сорта Шасла повышался на 40%, а Додреляби — на 70%. Слабое затенение не вызывало изменения внешнего вида ягод. Только при сильном затенении образование пигментов кожицы задерживалось и ягоды сорта Додреляби принимали слабо-розовую окраску вместо обычной черной.
В опытах Спаркса и Ларсена [492] при затенении кустов сорта Конкорд за 5—7 недель до уборки урожая на 30 и 50% содержание растворимых сухих веществ (РСВ) 1 в ягодах уменьшилось на 1—3%; общий сырой урожай с куста, вес ягод и количество ягод в грозди были на уровне контроля. По мере увеличения густоты кроны, числа побегов и урожая содержание РСВ в ягодах снижалось, причем это в большей мере зависело от плотности ягод, чем от соотношения между площадью листьев и урожаем гроздей.
В серии работ, выполненных Кливером с сотрудниками [441—444], с уменьшением облученности уменьшался размер ягод, снижалось содержание в них РСВ и пролина, задерживалось созревание. Одновременно в ягодах увеличивалось содержание общих кислот, яблочной кислоты, аргинина и общего азота. Спустя 24 ч после ввода С14 в побеги сорта Рислинг белый [445] в вариантах, где освещенность составляла 20 и 50 тыс. лк, количество яблочной кислоты становилось в 2—3 раза больше, чем при полной освещенности. Напротив, содержание винной кислоты было выше на полном свету. Интенсивность инсоляции не оказывала существенного влияния на содержание глюкозы и фруктозы.
По многолетним наблюдениям Вейнара [503] на шести сортах винограда (Мюллер-Тургау, Сильванер, Рислинг и др.), содержание винной кислоты в ягодах не зависит от температуры, продолжительности освещения и количества атмосферных осадков. Количество яблочной кислоты повышается при уменьшении температуры и продолжительности инсоляции. Положительная связь отмечается между содержанием сахаров в ягодах, длительностью солнечного освещения и температурой воздуха.
1 Растворимые сухие вещества (РСВ) — все растворимые компоненты соки ягод винограда неуглеводного характера.
Обстоятельная работа по выявлению роли света в окрашивании ягод винограда выполнена Уивером и Мак Кьюном [502]. Грозди сортов Токай, Султанина розовая и Эмперор в момент завязывания ягод изолировались от света и вплоть до уборки урожая находилась вне действия прямых солнечных лучей. Затенение практически не повлияло на созревание: среднее содержание РСВ и кислот в ягодах было соответственно 17,5 и 0,90% у растений на полном свету и 17,7 и 0,93% —у затененных. У сорта Токай при полном отсутствии света ягоды не окрасились. При частичном затенении грозди окрашивалась та ее часть, которая подвергалась действию солнечных лучей.
В другом опыте у сорока сортов винограда ягоды изолировались от света до начала их окрашивания. В момент уборки урожая у всех сортов, за исключением сортов Токай и Султанина розовая, ягоды затененных гроздей имели такую же окраску, как и на полном свету. Затенение не оказало влияния на содержание РСВ и кислотность ягод. По мнению авторов, действие света на окрашивание винограда зависит от пигментного состава ягод. В частности, для образования пигментов сорта Токай с Rf 0,23 и 0,14 свет необходим. Группа пигментов с Rf 0,28 и 0,10 может развиваться в темноте. Для большинства сортов действие света в период от завязывания ягод до начала окрашивания не было обязательным.
В исследованиях Наито [475, 476] многие сорта винограда со слабой окраской ягод обнаружили высокую требовательность к свету. В то же время группа сортов с темноокрашенной ягодой не имела различий по степени окраски ягод, если даже грозди созревали в темноте.
По данным Кливера [440], реакция сортов Кардинал и Пино гри на интенсивность света неодинакова. У сорта Кардинал при температуре 20° С и освещенности 5—12 тыс. лк ягоды окрашивались также или несколько интенсивнее, чем при той же температуре и освещенности 25—50 тыс. лк. У сорта Пино гри независимо от температуры воздуха ягоды окрашивались всегда слабее при низкой освещенности.
Таким образом, между освещенностью и качественным составом биопродуктов ягод обнаруживаются довольно сложные зависимости. В одном случае действие света на содержание биокомпонентов ягод проявляется отчетливо (содержание яблочной кислоты), в другом — менее ясно (содержание сахаров и IBC). Неоднозначно проявляется действие света и на содержании красящих веществ в ягодах. Сложности эти определяются, очевидно, тем, что хотя в экспериментах изучается действие света, его трудно вычленить из совокупного действия других факторов. А их может быть много: географическое местоположение виноградника, определяющее поступление суммарной солнечной радиации, сортовые особенности (особенно проявившиеся в окрашивании ягод), густота кроны, соотношение между листовой поверхностью и урожаем и др. Немаловажное значение имеет и то обстоятельство, что в опытах, проводимых в естественных условиях, обычно регулируется освещенность, но сохраняются естественные условия температурного режима воздуха, роль которого в формировании качества ягод показана всей многовековой практикой культуры винограда Нее это говорит о том, что для практических целей необходимо осуществлять работы для конкретизации действия светового фактора на качественный состав биопродуктов ягод в данной конкретной географической зоне на существующем сортовом материале, учитывая при этом, что качество ягод формируется и результате совокупного действия многих факторов, как внешних, так и обусловленных состоянием растения.