Все органические вещества в ягоде винограда подвержены значительным изменениям в процессе роста и созревания ягод — одни увеличиваются, другие уменьшаются. В большинстве случаев увеличение количества одних компонентов сопровождается уменьшением других, причем иногда процессы протекают синхронно в мякоти и кожице. Нередко, однако, они имеют противоположный характер. Раскрытие сути этих, порой довольно сложных, биохимических процессов является задачей изучения физиологических основ созревания винограда.
Изменение содержания сахаров и органических кислот
Из всех органических веществ, входящих в химический состав ягод, наиболее существенное значение для определения вкусовых и питательных качеств винограда и его пригодности к технической переработке имеют сахара и органические кислоты. Поэтому в большинстве случаев качество винограда определяют именно по содержанию сахаров и органических кислот, сколь бы узким и односторонним ни было это понимание.
После оплодотворения цветка и в начальные этапы роста завязей количество сахаров очень невелико, поскольку все ассимиляты используются на рост ягод. Более заметное увеличение содержания сахаров отмечается незадолго до начала созревания, а поворотный момент в накоплении сахаров в ягоде наступает в начале созревания. Все авторы, изучавшие динамику сахаров в процессе роста и созревания ягод винограда, отмечают, что в период начала созревания происходит как бы скачок в процессе сахаронакопления. В подтверждение сказанного выше отметим результаты исследований Стоева и Занкова (1958) относительно динамики накопления сахаров в винограде. Наряду с данными о накоплении сахара за один день на 1000 g винограда, на рис. 8 представлено уменьшение общей кислотности, вычисленное также за один день на 1000 g винограда.
Как видно, на начальных этапах роста ягод изменения в содержании сахаров и кислот незначительны. С наступлением периода начала созревания винограда, хотя это отмечается не вполне одновременно для всех сортов (для сорта Памид—12—21/VII, для сорта Мускат белый — 21/VII—1 /VIII, для Гибрида Х-10 — 21—26/VII), начинается интенсивное накопление сахара в винограде и соответствующее интенсивное снижение кислот. В это время накопление сахара достигает максимума — 5,08 до 5,60 g/dm3 глюкозы в день, а общая кислотность ежедневно уменьшается на 0,93—1,74 g/dm3 винной кислоты. Следовательно, наиболее значительные качественные изменения содержания сахаров и кислот в винограде наступают в течение 10 дней.
Аналогичный характер имеют данные сопоставления динамики роста ягод и темпов изменения содержания сахаров и кислот (Hawker, 1969).


Рис. 8. Динамика накопления сахаров ( в g глюкозы на 1000 g ягод винограда)(сверху) и динамика уменьшения общей кислотности (в g винной кислоты на 1000 g ягод винограда) (внизу)
Рис. 9. Сопоставление изменения содержания сахаров (2) и концентрации цитокинина (1) (μg KA x kg- ) в процессе роста и созревания ягод винограда (сорта Bachus)
Скорость накопления сахаров в °Ое; О — 100 — дни цветения; I—IV — фазы роста и созревания ягод
Весьма сходные результаты получили Георгиев и Янков (1960) при изучении динамики сахаров некоторых сортов.
По мнению Winkief (1962), в течение этого периода обогащение сахаром на несколько дней опережает темп роста ягод, на основании чего можно прийти к заключению, что подъем кривой роста может обусловливаться притоком сахара в ягоду.
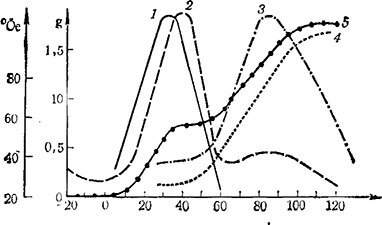
Рис. 10. Сопоставление изменений содержания ауксинов (1), цитокининов (2) и абсциссиновой кислоты (3) и накопления сахаров (4) в процессе роста и созревания ягод винограда
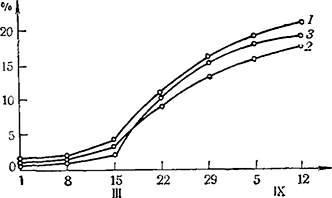
Рис. 11. Динамика накопления сахаров (%) в процессе созревания ягод винограда
1 — Алиготе; 2 — Рислинг; 3 — Каберне (по данным В. М. Лозы и В. В. Елецкого)
Роux (1950) удалось доказать, что максимальное содержание сахара в ягодах совпадает с моментом достижения максимального веса.
Вес ягод Bg, содержание сахаров в °бе, 0—120 — дни после цветения
Соотношение S/P (вес сахара ягод/вес ягод) в момент достижения их максимального веса колеблется незначительно по годам.
Coombe (1960) установил в начале созревания винограда сильно повышенное содержание ауксинов, что также может играть положительную роль в интенсивном накоплении сахаров. То же самое установлено и исследованиями Nitsсh (1953) и др.
Как установил Waitz (1975), в период максимального содержания цитокинина начинается накопление сахаров в ягодах, интенсивное же накопление их совпадает с понижением содержания цитокинина (рис. 9). В конце своей работы автор приводит суммарную схему, составленную Аllеwеldt, Waitz (1975), выражающую комплексное влияние ауксинов, цитокининов и абсциссиновой кислоты на рост ягод и накопление сахаров в них (рис. 10). Из данных видно, что начало интенсивного роста совпадает с максимальным содержанием ауксинов и цитокининов. Начало накопления сахаров не совпадает с резким снижением их содержания. Второй подъем роста ягод и интенсивное сахаронакопление приурочено к максимальному содержанию абсциссиновой кислоты.
Скачкообразный характер накопления сахаров в ягодах установлен и многими другими авторами. Козенко (1964), после математической обработки данных о динамике сахаров в винограде сортов Алиготе, Рислинг и Каберне Совиньон, полученных Лозой и Елецким (1933), приводит типичные кривые изменения концентрации сахаров (рис. 11). Эти кривые показывают, что существует ясно выраженная граница накопления сахаров между концом первого периода роста винограда и началом созревания. Как отмечает автор, на границе между двумя периодами наблюдается резкая активизация процесса накопления сахара.
Из более старых исследований, посвященных динамике сахаров и кислот в период роста и созревания винограда, следует упомянуть работы Mach и Роrtе1е (1880), Роrtе1е (1882) и Клоц (1933). На основании приводимых ими данных можно установить, что на протяжении полутора месяцев после оплодотворения цветков в растущих ягодах накопляется совершенно незначительное количество сахаров (см. Ribereau-Gayon, Peynaud, 1960). При вступлении ягод винограда в фазу начала созревания происходит интенсивное сахаронакопление, так что в течение 7—10 дней обнаруживается резкое изменение в содержании сахаров. После этого виноград вступает в фазу более равномерного сахаронакопления (табл. 5, 6).
Таблица 5
Динамика содержания сахаров и общего количества кислот в ягодах винограда
Время сбора | По Mach и Рortele (1880) | (сорт Неrрара, в 100 cm3 сусла, g) | По данным Роrtele (1882) (сорт Неrрара, g на 1000 ягод) | |
сахара, g | общее количество кислот, % | сахара, g | общая кислотность, | |
6/VII | 0,85 | 2,63 | 2,9 | 11,4 |
16/VII | 0,80 | 3,07 | 4,1 | 20,9 |
12/VIII | 1,96 | 3,36 | 16,4 | 34,0 |
23/VIII | 11,90 | 1,53 | 160,0 | 25,1 |
31/VHI | 13,40 | 1,06 | 217,0 | 21,2 |
29/1X | 17,10 | 0,85 | 312,0 | 15,4 |
12/Х | 21,20 | 0,65 | 382,0 | 14,8 |
Таблица 6
Динамика сахаров в созревающих ягодах винограда, g/dm3
(по Колоцу, 1933)
Время сбора | Сорт | Сорт | Сорт Шардоне |
7/VII | 0,90 | 1,10 | 0,95 |
14/VII | 3,00 | 4,20 | 3,80 |
21/VII период | 9,16 | 7,96 | 7,00 |
28/VII роста | 10,75 | 8,96 | 8,00 |
4/VIII ягод | 21,8 | 101,60 | 19,56 |
14/VIII | 79,60 | 130,00 | 83,50 |
18/VIII период | 129,00 | 163,20 | 103,20 |
25 /VIII созревания | 145,60 | 171,80 | 152,20 |
1/IX ягод | 214,00 | 222,60 | 210,20 |
Что же касается изменений общего количества кислот, то их динамика имеет совершенно противоположный характер: до начала созревания оно увеличивается, после чего наступает уменьшение, причем вначале более резкое, а затем равномерное. Аналогичное явление обнаружено Козенко (1962) при изучении динамики титруемой кислотности ягод.
Подобный характер имеют также данные о накоплении сахаров в винограде, установленные К о н д о (1944). Они весьма убедительно показывают, что все исследованные сорта (Дорой белый, Вассарга и Нимранг) в начале роста ягод (I период) содержат ничтожное количество сахара (0,28—0,51 %).
С наступлением начала созревания начинается интенсивное накопление сахара, о котором можно судить по крутому подъему кривой содержания сахаров. Кривые содержания кислот имеют вполне противоположный ход.
Данные Winkler (1962) также показывают, что в течение примерно 10 дней происходят резкие изменения в содержании сахаров и кислот у сортов Мускат александрийский и Аликант Буше.
Интенсивное сахаронакопление в ягодах винограда к началу созревания объясняется по-разному отдельными авторами. Moreau и Vinet (1932) считают, что большая часть сахаров поступает в ягоды из штамба и рукавов. Это мнение разделяет также Winkler (1962). "Такое передвижение, —говорит он, — часто приводит к быстрому накоплению сахаров в ягодах в начале их созревания".
Этого взгляда полностью придерживаются Ribereau-Gayon и Peynaud (1960). Рассматривая накопление сахаров в фазе начала созревания, они отмечают, что процессы эти длятся всего несколько дней, однако за это время происходит резкое изменение винограда в результате внезапного и обильного притока сахаров. В течение лишь одной недели, говорят они, количество сахаров в первых созревающих ягодах увеличивается в 6—7 раз.
Резкое накопление сахаров в период созревания Ribereau-Gayon и Peynaud объясняют притоком запасных веществ из побегов, многолетних частей, черешков листьев, листьев и гребней. В подтверждение этому авторы приводят данные исследования Moreau и Vinet (1932), из которых следует, что, по-видимому, существует уравновешенный баланс глюцидов (углеводов) в разных органах лозы (табл. 7).
Таблица 7
Баланс глюцидов (углеводов) в виноградной лозе (% к общему содержанию сахаров)
| Органы лозы | До начала созревания | После начала созревания |
Тонкие корешки | 2,6 | 3,8 |
Толстые корни | 12,0 | 13,4 |
Ствол и его ветвления | 27,4 | 18,1 |
Зеленые побеги | 21,4 | 14,6 |
Черешки листьев | 2,2 | 1,7 |
Пластинки листьев | 17,7 | 12,6 |
Гребни | 1,3 | 1,1 |
Ягоды | 15,2 | 34,3 |
Несколько иную точку зрения выражают Lafon-Lafourcade и Peynaud (1959). По их мнению, сахара и крахмал, которые к периоду ослабления роста побегов накопились в листьях и побегах, перемещаются в ягоды (они в это время достигают 3/4 своего максимального размера). Почти аналогичное мнение высказывает Marteau (1955), который отмечает, что во время созревания интенсивность миграции сахаров к ягодам обусловлена, видимо, уменьшением резервных глюцидов, накопленных в листьях, иными словами — ассимиляцией хлорофилла.
Сисакян с сотр. (1948), изучая превращение углеводов в листьях, гребнях и ягодах нескольких сортов, обнаружили четкую зависимость между уменьшением углеводов в листовых пластинках и возрастанием их количества в ягодах в фазе начала созревания ягод. В результате мобилизации веществ из листьев в проводящие и запасные органы, в процессе активного роста и созревания ягод в листовых пластинках имеет место резкое падение содержания как подвижных, так и запасных форм углеводов.
Мнение о перемещении запасных углеводов из многолетних частей к ягодам к периоду начала созревания нельзя считать правдоподобным. По данным Стоева (1947—1948), корни отправляют углеводы из накопленных в конце предыдущего вегетационного периода резервов примерно до конца июня, а многолетние части куста — намного раньше. Кроме того, невозможно делать заключение о перемещении углеводов из многолетних частей лишь на основании данных об уменьшении количества сахаров в них. Как показали исследования Стоева (1947—1948) и исследования ряда других авторов, в период начала созревания происходит не только резкое накопление сахаров в ягодах винограда, но и резкое усиление синтеза веществ, в результате чего наблюдаются интенсивное превращение сахаров в крахмал и накопление крахмала в древесных частях, интенсивное вызревание побегов и другие процессы, что свидетельствует о качественном изменении всего обмена веществ. Поэтому правильнее предположить, что интенсивное накопление сахара в ягодах к периоду начала созревания имеет ритмический характер и является качественно переломным моментом в годичном цикле развития виноградной лозы (Стоев, Занков, 1958). Следует добавить, что согласно исследованиям Stoev и Ivantschev (1977) в период начала созревания ягод отток ассимилятов к многолетним частям и корням винограда временно (на 8-10 дней) приостанавливается и все продукты фотосинтеза листьев направляются к ягодам винограда*. Это создает предпосылки для тотального передвижения ассимилятов и спонтанного их накопления в ягодах в период начала созревания, что дает нам основание назвать это явление феноменом veraison.
Ряд других авторов изучал также изменения содержания сахаров и органических кислот в процессе созревания ягод винограда (Маrtеаu, 1955; Герасимова, Дихтяр, 1953; Патарая, Силагадзе, 1962; Кондо, 1944, 1946; Марутян, 1956, 1957; Журавель, 1960; Кондо, Короткевич, 1959; Кананадзе, 1950; Peynaud, 1946; Берг, 1948; Иванов, 1963; Милованова с сотр., 1960; Георгиев, 1938; Dеibnеr, 1955; Кравченко, 1972 и др.). Множество данных о динамике сахаров и кислот во время созревания ягод имеется и в работах Берг (1940), Фролова-Багреева, Агабальянца (1951), в Ампелографии СССР (1946—1956), Ampelografia Republici Socialista Romania (1959— 1967), Ribereau-Gayon и Peynaud (1960) и др., которые не представляется возможным рассматривать в отдельности. В большей части этих работ, в особенности там, где исследования содержания сахаров и кислот в ягодах винограда проводились в динамике, отмечается поворотный момент в накоплении сахаров в фазе начала созревания или же уменьшение кислот, что соответствует указанной выше закономерности.
Многочисленные исследования проводились и в целях установления природы сахаров и органических кислот, накопляющихся в винограде во время его созревания.
Почти все авторы, изучавшие вопрос о компонентах сахара, показали, что на начальном этапе в ягодах винограда преобладает глюкоза, а фруктоза находится в минимальном количестве — в виде следов. После начала созревания винограда в ягодах накапливаются и глюкоза и фруктоза, причем, по данным Кондо (1944), интенсивнее происходит накопление фруктозы. Например, у раннеспелого сорта Ак-халили количество фруктозы увеличилось в 40 раз (с 0,06 до 2,4%), в то время как количество глюкозы увеличилось только в 5 раз (с 0,48 до 2,65 %). Аналогичная зависимость установлена и у сорта Дорой белый.
У поздних сортов (Нимранг и Тайфи) после наступления момента интенсивного накопления сахаров фруктоза увеличивается более быстрыми темпами по сравнению с глюкозой, хотя соотношение между количеством фруктозы и глюкозы не столь резкое, как у ранних сортов. Как отмечает Кондо (1944), однако, к концу второй фазы (третий период) фруктоза явно преобладает над глюкозой.
Неравномерный темп накопления глюкозы и фруктозы в винограде в различные этапы его формирования и созревания подтверждается и исследованиями Марутян (1956), Ribereau-Gayon и Peynaud и др.
Winkler (1962) считает, что глюкоза преобладает главным образом во время роста ягод, тогда как после созревания количество глюкозы и фруктозы почти одинаково. Если виноград останется несозревшим, основным сахаристым веществом будет глюкоза. По мнению Миловановой (1960), повышение относительной влажности воздуха приводит к понижению содержания фруктозы.
Г е т о в (1966а) почти всегда устанавливал значительное превалирование фруктозы (в 1,5 раза) у сильно перезревших ягод. Повышение содержания фруктозы при перезревании винограда и длительном его хранении и в результате этого значительное повышение соотношения фруктоза: глюкоза наблюдали также Venezia и Gentilini (1935).
При подробном изучении содержания и форм сахара в винограде 26 сортов — армянских, грузинских, азербайджанских, венгерских, французских и американских, Сисакян и Марутян (1948) установили, что не существует строго определенной закономерности в накоплении глюкозы и фруктозы. Они считают, что соотношение глюкоза:фруктоза в процессе созревания подвергается значительным колебаниям с переменным преобладанием одного или другого моносахарида. У некоторых сортов в физиологической зрелости преобладает фруктоза по сравнению с глюкозой, у других—наоборот.
Вопрос о содержании глюкозы и фруктозы в винограде и, в особенности, об их соотношении, очевидно, не вполне решен, так как в литературе встречаются значительные противоречия. Genevois и Ribereau-Gayon (1947) считают, что оба сахаристые вещества находятся в спелом винограде в эквимолекулярных количествах. Почти в одинаковых количествах содержатся глюкоза и фруктоза и, по данным Peynaud (1947), соотношение Г:Ф колебалось от 0,93 до 0,96 в 1937 г. и от 0,91 до 0,94 в 1938 г. Коробкина установила, что в полной зрелости винограда фруктоза слегка преобладает по сравнению с глюкозой.
Таблица 8
Содержание глюкозы и фруктозы и соотношение между ними у различных сортов
Сорт | Дата сбора урожая | Содержание сахаров, % | Глюкоза, g/100 cm3 | Фруктоза, g/100 cm8 | Глюкоза/ |
Столовые сорта | |||||
Alphonse Lavallee | 13/X | 20,0 | 9,74 | 9,87 | 0,99 |
Cardinal | 16/VIII | 20,0 | 10,23 | 9,30 | 1,10 |
Chasselas doree | 24/IX | 20,0 | 9,87 | 9,80 | 1,01 |
Flame Tokay | 20/IX | 20,0 | 9,12 | 10,81 | 0,84 |
Khalili | 30/VIII | 14,4 | 6,10 | 6,01 | 1,01 |
Muscat of Alexandria | 3/IX | 20,6 | 9,2] | 11,12 | 0,83 |
Red Malaga | 26/IX | 20,0 | 11,03 | 9,84 | 1,12 |
Thompson Seedless | 3/IX | 22,3 | 10,36 | 12,94 | 0,80 |
Винные сорта | |||||
Cabernet Sauvignon | 3/IX | 21,0 | 9,44 | 9,69 | 0,98 |
Delavare | 3/IX | 21,8 | 9,95 | 10,87 | 0,92 |
Gewurzttraminer | 3/IX | 22,3 | 10,19 | 10,78 | 0,95 |
Folle Blanche | 16/X | 18,3 | 8,31 | 9,22 | 0,90 |
Gamay Beaujolais | 22/VIII | 20,0 | 9,47 | 8,86 | 1,07 |
Mission | 23/IX | 22,2 | 10,10 | 0,81 | |
Palomino | 23/X | 16,4 | 6,32 | 7,94 | 0,80 |
Sauvignon blanc | 3/IX | 20,0 | 8,97 | 10,12 | 0,89 |
Sylvaner | 3/IX | 21,1 | 9,38 | 10,95 | 0,86 |
Valdepenas | 3/IX | 20,8 | 10,19 | 12,22 | 0,83 |
White Riesling | 3/IX | 21,2 | 9,63 | 11,54 | 0,83 |
По данным Bayer (1958), соотношение глюкоза: фруктоза колеблется между 0,83 и 1,21. Amerine и Thookis (1958), подтверждая взгляд, что количество глюкозы и фруктозы зависит от степени созревания винограда, установили, что соотношение между ними неодинаково для винных и столовых сортов — фруктоза в большей степени преобладает у винных сортов (табл. 8).
Соотношение между глюкозой и фруктозой в винограде имеет не только теоретическое, но и практическое значение. Как известно, фруктоза значительно слаще других сахаров — 15 %-ный раствор фруктозы дает такое же ощущение сладости, как 22,8 %-ный раствор глюкозы или 17,8 %-ный раствор сахарозы. Поэтому подбор форм, в которых преобладает фруктоза, приобретает значительное практическое значение в селекционной работе при выведении ранних столовых сортов или сортов, предназначенных для выращивания в более северных районах, как и для получения виноградного сока. Кроме моносахаридов, в винограде накопляется известное количество сахарозы.
Вопрос о наличии сахарозы в ягодах винограда исследовался в ряде работ. В более старых работах отмечалось, что у европейских сортов имеются лишь следы сахарозы в винограде. В американских же сортах установлено значительное количество сахарозы (Thompson, Whitter, 1913; Ваragiola, Godet, 1914; Gore, 1909; Gardwell, 1925). Сахароза имеется и в ягодах мичуринских сортов (Франчук, 1934), а также в среднеазиатских сортах винограда (Кондо, 1944). Сисакян и Марутян (1948) установили наличие сахарозы в сортах винограда, относящихся к различным эколого-географическим группам. В число изученных сортов вошли армянские, грузинские, азербайджанские, венгерские, французские и американские сорта.
Стоев с сотр. (1960) изучали содержание сахарозы в ягодах 30 сортов винограда различного происхождения. При этом определение сахаров проводили дважды: в начале созревания и в физиологической зрелости ягод.
Полученные результаты показали, что в винограде большинства сортов в начале созревания обнаруживаются лишь фруктоза и глюкоза, хотя в это время в ножках ягоды имелась сахароза. Это дает основание авторам предполагать, что ассимиляты, выработанные в листьях, передвигаются вниз по нерватуре и черешкам листьев, по побегам, гребням кистей и ножкам ягод в форме сахарозы. Затем на начальных этапах роста ягод (до начала созревания) она не обнаруживается в ягодах винограда.
В период физиологической зрелости ягод сахароза обнаруживалась в ягодах всех исследовавшихся сортов, относящихся ко всем экологическим группам: понтийской, западноевропейской, среднеазиатской и к группе прямых производителей. Это позволяет считать, что наличие сахарозы в ягодах в период их физиологической зрелости характерно для виноградной лозы и не зависит ни от сортовых особенностей, ни от происхождения сорта — появление сахарозы в ягодах винограда приурочено к моменту общего повышения в них содержания сахаров — к III периоду роста ягоды.
Hardy (1968) также установил, что меченая по углероду сахароза переходит в радиоактивную глюкозу и фруктозу под воздействием инвертазы. На этом основании автор считает, что путь поступления сахара через цветоножки в ягоды может быть иным, чем канал передвижения ассимилятов из листа, или же что поступающая в ягоды сахароза подвергается гидролизу.
Можно указать еще ряд работ, в которых рассматривается вопрос о наличии сахарозы в ягодах (Кондо, 1944 и др.). Коробкина (1961) из изучаемых 6 столовых сортов не установила сахарозы лишь в двух (Нимранг и Тайфи розовый). Патарая и Силагадзе (1962), полностью подтверждая наши данные (Стоев с сотр., 1960), что сахароза отсутствует в начальные этапы созревания винограда, а появляется позже, в период технической зрелости у всех сортов, выражают мнение, что сахарозу можно считать объективным показателем физиологической зрелости винограда.
Ribereau-Gayonn Peynaud (1960), однако, считают, что в ягодах имеются лишь едва заметные следы сахарозы. По их мнению, при выходе из листа сахароза в сосудах гидролизуется и количество ее по пути от черешков листьев до гребней и ягод винограда непрерывно уменьшается.
Более вероятным является предположение, что гидролиз сахарозы происходит при ее поступлении в ягоды винограда, причем это отмечается до периода начала созревания, чему, по-видимому, благоприятствует кислотный баланс ягоды. Это предположение лучше согласуется с установившимся в науке положением о роли сахарозы как основного средства транспорта для растений и, в частности, для виноградной лозы (Соунсон, 1961; Чанишвили, 1964; Курсанов, 1976).
При рассмотрении вопроса о сахарозе в ягодах винограда заслуживает внимания сообщение Марутян (1956), что в ягодах содержится активный фермент инвертаза. По мнению Аrnоu1d (1967), инверсия сахарозы до глюкозы и фруктозы в созревающих ягодах винограда происходит под влиянием инвертазы.
По данным Марутян (1956), сахароза увеличивается в винограде после ранних осенних заморозков, что, очевидно, происходит за счет уменьшения моносахаридов.
Исследования Hardy (1967), проводимые с применением меченого источника углерода, дают возможность судить о метаболизме сахаров в ягоде винограда. С этой целью он помещал ягоды сорта Султанина (Кишмиш белый) с ножкой длиной в 5 мм в специальную аппаратуру, заполненную дистиллированной водой, насыщенной радиоактивными сахарами (14С сахароза, 14С фруктоза и 14С глюкоза). После 7,5 и 24-часовой экспозиции автор удалял ножки ягод и подвергал их обработке по соответствующей методике, которая позволяла делать заключения об изменениях, происходящих с поглощением ягодой радиоактивных сахаров. Средний вес ягоды к моменту эксперимента составлял 1,106+0,10 g; средняя концентрация глюкозы и фруктозы— соответственно 3,34+0,32 и 3,31+0,31 g в пересчете на 100 g свежего веса. Таким образом, развитие ягоды находилось в фазе интенсивного накопления сахаров и интенсивного уменьшения содержания кислот.
Результаты исследования показали, что после 7,5-часовой экспозиции радиоактивность сахаров и органических кислот в обработанных 14С сахарозой ягодах составляет около 65 % общего количества вносимой радиоактивности, в то время как радиоактивность ягод, обработанных 14С глюкозой и 14С фруктозой, была значительно ниже. На этом основании автор приходит к выводу, что сахароза подвергается метаболизму в большей степени, чем глюкоза и фруктоза. Из последних двух сахаров (глюкоза и фруктоза) глюкоза показывает более интенсивный метаболизм. После 24-часовой экспозиции отмечалось уменьшение радиоактивности ягод, обработанных 14С сахарозой и 14С фруктозой.
Особо следует отметить констатацию автора, что при экзогенном введении 14С глюкозы и 14С фруктозы в ягоды обнаруживается механизм внутреннего метаболизма этих сахаров, причем синтез сахарозы установлен во всех ягодах, в которые вводились радиоактивные моносахариды.
Синтез сахарозы из экзогенно введенных моносахаридов указывает также на наличие активности одного или двух ферментов, синтезирующих сахарозу или сахарозофосфаты: UDP*-глюкозо-фруктозо-транс-глюкозидаза или UDP*- глюкозо-фруктозо-6-фосфат-транс-глюкозидаза. Таким образом, синтез сахарозы в ягодах винограда можно представить следующим образом:
UDP-глюкоза+фруктоза↔UDP+сахароза.
*UDP — уридин-дифосфат.
Более поздние исследования Hardy (1968) дают некоторые дополнительные сведения о метаболизме сахаров в ягодах винограда. Установлено, что вводимый радиоактивный углерод 14С в форме равномерно меченой сахарозы в ножках ягод лишь в 2С—25% остается неизменным, а 50—60% переходят в глюкозу и фруктозу. Следовательно, сахароза сразу после ее поступления в ягоды переходит в низкомолекулярные соединения. В последующие 12 h количество радиоактивной сахарозы несколько уменьшается. Характерно еще то, что количество глюкозы и фруктозы неодинаково, а потому соотношение между ними не равняется единице — фруктозы больше, чем глюкозы.
При введении радиоактивной сахарозы, меченой лишь в фруктозной части, соотношение 14С глюкоза/14С фруктоза на протяжении 12 h доходит до единицы, из чего автор делает вывод, что сначала произошел распад радиоактивной сахарозы, а позже ресинтез. Hardy считает также, что радиоактивная глюкоза, продукт равномерно меченой сахарозы, образуется из глюкозной части сахарозы, а не путем превращения фруктозы в глюкозу в результате распада сахарозы на UDP-глюкозу и фруктозу.
Кроме того, автор приходит к заключению о невозможности UDP-глюкозы быть посредником в образовании глюкозы из сахарозы по схеме
UDP-глюкоза![]() Hardy приходит к этому выводу на основании неустойчивости неорганического фосфата (Мurata с сотр., 1966), и содержания пирофосфатов в растениях в значительных количествах (Long, 1961). Поэтому считается, что реакция UDP-глюкозопирофосфорилаза протекает преимущественно в одном направлении — в сторону синтеза UDP-глюкозы (Мuratа с сотр., 1966).
Hardy приходит к этому выводу на основании неустойчивости неорганического фосфата (Мurata с сотр., 1966), и содержания пирофосфатов в растениях в значительных количествах (Long, 1961). Поэтому считается, что реакция UDP-глюкозопирофосфорилаза протекает преимущественно в одном направлении — в сторону синтеза UDP-глюкозы (Мuratа с сотр., 1966).
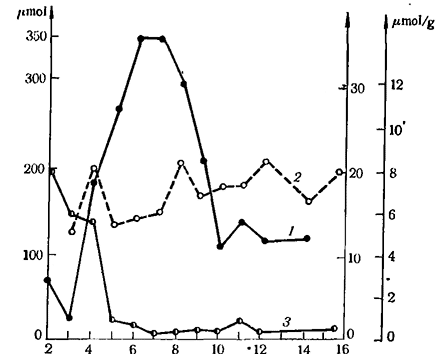
Рис. 12. Активность инвертазы (μmol сахарозы) (1), гексокиназы (в уcл. ед. на g свежею веса) (2) и глюкозо-6-фосфатдегидрогеназы (μmol на g свежего веса за час) (3) в ягодах сорта Султанина; 2—16—недели после цветения
Предполагается, что преимущественное образование фруктозы при введении равномерно меченой сахарозы в ножки ягод является результатом ускоренного проникновения глюкозы после распада сахарозы в участки активного метаболизма. Наоборот, при введении в листья винограда меченой углекислоты 14СО2 из ассимилятов доминирует радиоактивная глюкоза (Кliеwеr, 1964; Кliеwеr, Schultz, 1964).
Об участии ферментов в метаболизме сахаров в ягодах винограда сообщает Hawker (1969). В течение недели в период развития ягод винограда сорта Султанина исследовались изменения активности инвертазы, гексониназы, глюкозо-6-фосфатгидрогеназы, синтетазы сахарозы, фосфатной синтетазы сахарозы и фосфатазы сахарозы в экстрактах, обработанных карбоваксом 4000. Отмеченному выше быстрому накоплению сахаров в ягодах в середине процесса их развития, т. е. в начале созревания (см. рис. 6, 7), предшествовало усиление активности инвертазы (рис. 12). Почти одновременно усиливалась активность синтетазы сахарозы, фосфатной синтетазы сахарозы и фосфатазы сахарозы. Активность глюкозо-6-фосфатдегидрогеназы уменьшалась рано и оставалась на низком уровне в течение развития ягод. Предполагается, что инвертаза в ягодах винограда находится в цитоплазме или же в вакуолях клетки (Hawker, 1969).
Существенным компонентом ягоды винограда являются органические кислоты, содержание которых определяет общую титруемую кислотность. Как отмечалось, общая титруемая кислотность подобно сахарам подвергаться весьма значительным изменениям, играет важную роль в процессе созревания ягод и в определении качества сусла и получаемых вин. Поэтому вопрос о содержании органических кислот и об их изменениях в период созревания ягод винограда всегда вызывал значительный интерес среди исследователей физиологии винограда и технологов виноделия.
В ягоде виноградной лозы содержатся винная, яблочная, лимонная, аскорбиновая, фосфорная кислоты, а в небольших количествах и другие органические кислоты. Винная и яблочная кислоты составляют свыше 90% общих кислот (Winkler, 1962). Доля остальных кислот в ягодах винограда незначительна (Colagrande, 1959).
Родопуло (1960) установил щавелевую, винную, лимонную, яблочную, гликолевую и янтарную кислоты в сусле разных сортов винограда. Почти аналогичный состав органических кислот установили Журавель (1960), Фролов-Багреев и Агабальянц (1951), Иванов (1963) и др.
Георгиев и Янков (1959, 1960) утверждают, что до начала созревания винограда установили только винную и яблочную кислоты. Изучением винной и яблочной кислот в начале ограничилась Никова (1962), однако позже она установила еще лимонную и летучие кислоты (1966).
Некоторые авторы (Alquier-Boufard, Carles, 1963; Carles, Alquier-Boufard, 1962) изучали органические кислоты почти во всех органах виноградной лозы. В ягодах винограда, кроме винной и яблочной, они установили еще лимонную, янтарную, фумаровую, глицериновую, хининовую и шикимовую кислоты. Они представлены в небольших количествах и не всегда обнаруживаются в ягодах винограда.
В литературе еще не получил полного выяснения вопрос о происхождении основных органических кислот в ягоде винограда. Некоторые авторы (Stafford, Loewns, 1958; Stafford, 1959) считают, что синтез винной кислоты происходит в листьях винограда, откуда она переходит в ягоды (Genevо s с сотр., 1954; Gadet, 1939). Другие авторы, наоборот, указывают, что синтез винной и яблочной кислот в основном совершается в зеленой ягоде (Halе, 1962, 1962а).
Кliеwеr (1964) и Кliеwеr, Schultz (1964) установили, что после введения 14СO2 в листья меченая яблочная кислота синтезировалась быстрее, чем винная. Установлено также, что синтез меченых форм яблочной и винной кислот происходит быстрее в незрелых ягодах, чем в созревших. Однако в этих исследованиях оставался невыясненным вопрос, передвигаются ли органические кислоты из листьев в меченой форме или же образуются в ягодах из радиоактивной сахарозы.
Некоторое выяснение этот вопрос получил в работе Hardy (1968), установившего, что при введении в ножки ягод радиоактивной сахарозы наблюдалось быстрое превращение сахарозы в яблочную и винную кислоты: за 3 h 15% радиоактивного углерода, введенного через сахарозу в ягоды, превращались в яблочную кислоту. После 24 h обнаружено, что 7% радиоактивного углерода перешли в винную кислоту. Не установлено с достоверностью участия глюкозной и фруктозной частей сахарозы в синтезе органических кислот.
Переход меченого углерода из радиоактивной сахарозы в аминокислоты и другие органические кислоты (лимонную, гликолевую, янтарную и фумаровую) незначителен, и доля его несколько больше в начале экспозиции.
При введении 14С в форме глюкозы и фруктозы, L-яблочной кислоты и Dl-винной кислоты (1,4 14С) установлено, что метаболизм глюкозы осуществляется быстрее, чем 14С фруктозы. Установлено также, что и фруктоза, и глюкоза переходят в органические кислоты. Через 24 h после их введения в ножки ягод радиоактивные формы яблочной и винной кислот включали 25, соответственно 10% общей радиоактивности глюкозы и фруктозы. При введении L-яблочной кислоты основная часть ее радиоактивности приходится на гликолевую и янтарную кислоты.
Метаболизм винной кислоты происходит очень медленно — небольшие количества радиоактивного углерода переходят в гликолевую и яблочную кислоты, основная же часть (90%) спустя 24 h остается в винной кислоте.
Исследования динамики органических кислот в ягоде винограда показали, что по мере роста и созревания ягод главнейшие органические кислоты (яблочная и винная) претерпевают значительные изменения. По данным Миловановой с сотр. (1960), на начальном этапе роста ягоды (при сахаристости 2 %) основной органической кислотой является яблочная — содержание винной кислоты находится в пределах 7—12,9 g/dm3, в то время как содержание яблочной достигает 25—29,2 g/dm3. В дальнейшем, однако, снижение яблочной и винной кислот происходит неодинаковыми темпами — значительно быстрее уменьшается яблочная кислота. По достижении 18% сахаристости содержание яблочной кислоты уменьшалось в 8,5—13,3 раза, в то время как содержание винной кислоты снизилось всего в 1,3—2,3 раза. Суточное снижение винной кислоты составляет 0,14—0,19 g/dm3, для яблочной кислоты оно происходит главным образом за счет окисления, а для винной — за счет образования солей и повышения влажности ягод.
Неодинаковый темп уменьшения яблочной и винной кислот констатировали также Ribereau-Gayon и Peynaud (1960). После начала созревания содержание винной кислоты стабилизируется и мало изменяется. Но яблочная кислота продолжает уменьшаться и после наступления начала созревания довольно быстрыми темпами.
В связи с этим доля винной кислоты, которая на начальных этапах созревания (при сахаристости менее 10%) составляет около 35—40% общего количества винной+яблочной кислот, постепенно увеличивается и к периоду физиологической зрелости достигает 80—90%. При этом отдельные сорта ведут себя по-разному. Сорт Рислинг, например, теряет почти целиком яблочную кислоту, в то время как сорта Тербаш и Мускат венгерский сохраняют яблочную и винную кислоту в одинаковых пропорциях (Кондо, Короткевич, 1959).
Laub (1971) показал, что изменения содержания яблочной кислоты связаны с изменениями общей кислотности и содержания сахаров. Наиболее значительные изменения наступают при интенсивной солнечной радиации и при отсутствии осадков.
По мере созревания винограда уменьшается и количество других кислот — винной, яблочной, янтарной, щавелевой, пировиноградной (Родопуло, 1960).
Здесь следует отметить уникальный факт, о котором сообщают Воubals, Bourzeix и Guitraud (1977). Они установили, что у иранского сорта Gora Chenine содержание органических кислот очень низкое, причем яблочная и винная кислоты в процессе созревания винограда остаются почти в неизменном количестве.
Hawker (1969) сообщает, что в период активного снижения содержания яблочной кислоты в ягодах сорта Султанина не установлено значительного повышения активности яблочного энзима и пируват декарбоксилазы. Активность фосфопируват карбоксилазы, высокая в начале развития ягод, снижалась почти одновременно с приостановлением синтеза яблочной кислоты.
Сисакян с сотр. (1948) показали, что в период начала созревания наступает не только уменьшение кислотности ягод винограда, но также титруемой и действительной кислотности в листьях различной формации и в гребнях винограда. На основе полученных результатов авторы приводят к выводу, что в фазе начала созревания и полной физиологической зрелости в кислотообразующей способности растения наступает коренной сдвиг. "Это возникает, — отмечают авторы, — по-видимому, в результате перехода растения от кислотообразования к преимущественному сахаронакоплению".
Значительные изменения происходят также в действительной кислотности винограда. Как показали исследования Берг (1948), Миловановой с сотр. (1960) и др., в процессе созревания винограда параллельно со снижением титруемой кислотности происходит заметное повышение величины pH, причем у разных сортов это происходит с неодинаковой интенсивностью. У некоторых сортов к периоду технической зрелости винограда кривая динамики pH после крутого подъема занимает почти горизонтальное положение. У других же сортов и после достижения периода технической зрелости кривая pH идет по восходящей линии, что указывает на возможность дальнейшего снижения действительной кислотности.
Изучение динамики pH у разных сортов определяет срок сбора урожая с учетом биологических особенностей сортов и требований, предъявляемых технологией.
В ходе созревания винограда происходят изменения и т. наз. окислительновосстановительного потенциала (ОВ), выражающего степень напряженности процессов окисления. На начальном этапе созревания винограда ОВ-потенциал, обозначаемый Eh, достигает высокого значения, после чего закономерно снижается. Динамика Eh неодинакова у различных сортов, что может служить для технологической характеристики сортов и определения срока сбора урожая в зависимости от направления переработки.
Dеibnеr (1955) установил, что наиболее высокой напряженности ОВ- потенциал достигает к периоду начала созревания (табл. 9), т.е. сахаронакопление и снижение кислотности ягод винограда наиболее интенсивны.
После максимума окислительно-восстановительный потенциал прогрессивно падает и ко времени сбора урожая достигает довольно стабильной величины. Берг (1953а) отмечает, что к моменту технологической зрелости сортов для столовых вин ОВ-потенциал снижается до 306—374 mV.
Для более точной характеристики кислотности винограда применяется т. наз. ацидиметрический баланс. В нем сопоставляются катионы и анионы в процессе созревания винограда, что дает представление о динамике и изменениях всех компонентов кислотности винограда. В качестве примера в табл. 10 приводятся данные ацидиметрического баланса сорта Мальбек по данным Peynaud (1946). Многие такие балансы приведены в книге Ribereau-Gayon и Peynaud (1960).