Глава 10
БРОЖЕНИЕ ВИНОГРАДНОГО СУСЛА
Первые попытки вскрыть природу химизма образования этилового спирта из сахара были предприняты в начале XIX в. К этому времени А. Лавуазье и Л. Гей-Люсак сформулировали суммарное уравнение, основываясь на определении спирта и углекислоты:
![]()
Однако в то время существовали различные мнения по вопросу, какие реакции происходят при превращении сахара в спирт и углекислоту. В связи с этим в середине XIX в. между Ю. Либихом и Л. Пастером возник спор, в котором были затронуты основные философский проблемы биологии.
Ю. Либих, выдвинул химическую теорию процесса брожения, вызываемую белковыми веществами дрожжей, которые, разлагаясь, приводят к молекулярным колебаниям, при этом сахар распадается на спирт и углекислоту.

Одновременно Л. Пастер убедительными опытами доказал, что сбраживание сахара происходит только в присутствии микроорганизмов, являющихся возбудителями алкогольного брожения, так называемых дрожжей. Этим Л. Пастер сумел установить живую природу дрожжей и обосновать взгляд на брожение как на результат жизнедеятельности дрожжей и доказать несостоятельность теории Ю. Либиха.
Л. Пастер считал, что дрожжи, способные существовать без кислорода, приобретают энергию из сахара при разложении его на спирт и углекислоту.
Способность дрожжей к анаэробному брожению соответствует выходу спирта согласно уравнению разложения сахара на этанол и углекислоту. Но если их культивировать в аэробных условиях, то выход спирта уменьшается, так как часть сахара расходуется на дыхание.
Впервые Л. Пастер делает вывод, что выход спирта по отношению к израсходованному сахару значительно ниже.
Угнетение брожения дыханием О. Варбург назвал пастеровским эффектом.
Хотя некоторые исследователи выступили с критикой в отношении эффекта Пастера, но все же было установлено, что кислород благоприятствует размножению дрожжей, при этом выход спирта значительно уменьшается.
В аэробных условиях дрожжи переключаются с брожения на дыхание.
Это оправдалось в опытах С. П. Костычева, который установил, что в присутствии кислорода дрожжи не прекращают брожения и сбраживают 2/з сахара с одновременным его окислением на треть.
Для выявления механизма эффекта Пастера были проведены многочисленные исследования. Считают, что эффект Пастера является результатом взаимодействия между различными энергетическими путями, существующими у дрожжей. Одним из проявлений такого взаимодействия является концентрация аденозиндифосфата и неорганического фосфата между процессами субстратного фосфорилирования гликолитического пути и окислительного фосфорилирования в дыхательной цепи.
Л. Пастер установил связь между жизнедеятельностью дрожжей и процессом брожения, но какова была эта связь и как происходит процесс брожения, на это он ответить не смог.
Пролить свет на эти жизненно важные вопросы удалось только Э. Бюхнеру, который впервые показал, что спиртовое брожение является энзиматическим процессом.
Для выделения энзимов брожения Бухнер применил способ механического разрушения живых клеток под высоким давлением.
Исследования показали, что дрожжевой сок, полеченный при прессовании дрожжей, кроме зимазы содержит еще ряд ферментов.
Впоследствии было выяснено, что зимаза является смесью энзимов.
В результате многочисленных биохимических исследований было установлено, что процесс превращения глюкозы в этанол и углекислоту очень сложный и протекает через ряд промежуточных соединений с участием различных энзимов.
В этих работах принимали участие такие выдающиеся ученые, как А. Н. Лебедев, С. П. Костычев, К. Нейберг, Г. Эмбден, О. Мейергоф и др., которые выявили отдельные этапы спиртового брожения.
Химизм алкогольного брожения
В дрожжевой клетке содержится множество ферментов, которые осуществляют разные биохимические процессы, в том числе и алкогольное брожение.
Подготовительный этап брожения углеводов заключается в образовании фосфорных эфиров гексоз. Первые шаги в этом направлении были сделаны советскими учеными Л. А. Ивановым и Н. Лебедевым, которые обнаружили участие фосфатов в синтезе фосфорных гексоз.
Затем это подтвердили зарубежные исследователи А. Гарден и B. Юнг.
Исследования Л. А. Иванова и А. Н. Лебедева показали, что если к свежему дрожжевому соку добавить глюкозу, то скоро начинается брожение, которое затем постепенно замедляется, но которое можно восстановить прибавлением неорганического фосфора.
Выделенный ими из бродящей среды фосфорный эфир оказался фруктофуранозо-1,6-дифосфатом. Более глубокие Исследования этих ученых показали, что фосфорные эфиры гексоз состоят из равновесной смеси глюкопиранозо-6-фосфата и фруктофуранозо-6- фосфата.
Фосфолирование глюкозы является необходимым процессом для перевода ее в лабильное метаболитически активное соединение.
Действием фермента гексокиназы происходит перенос одного остатка фосфорной кислоты от аденозидтрифосфата (АТФ) на глюкозу, при этом образуется глюкопиранозо-6-фосфат и аденозиндифосфат (АДФ). Затем действием фермента гексозофосфатизомеразы глюкопиранозо-6-фосфат превращается в фруктофураназо-6-фосфат. Все эти реакции образования фосфорных эфиров гексоз на первой стадии брожения изображены в следующей схеме:
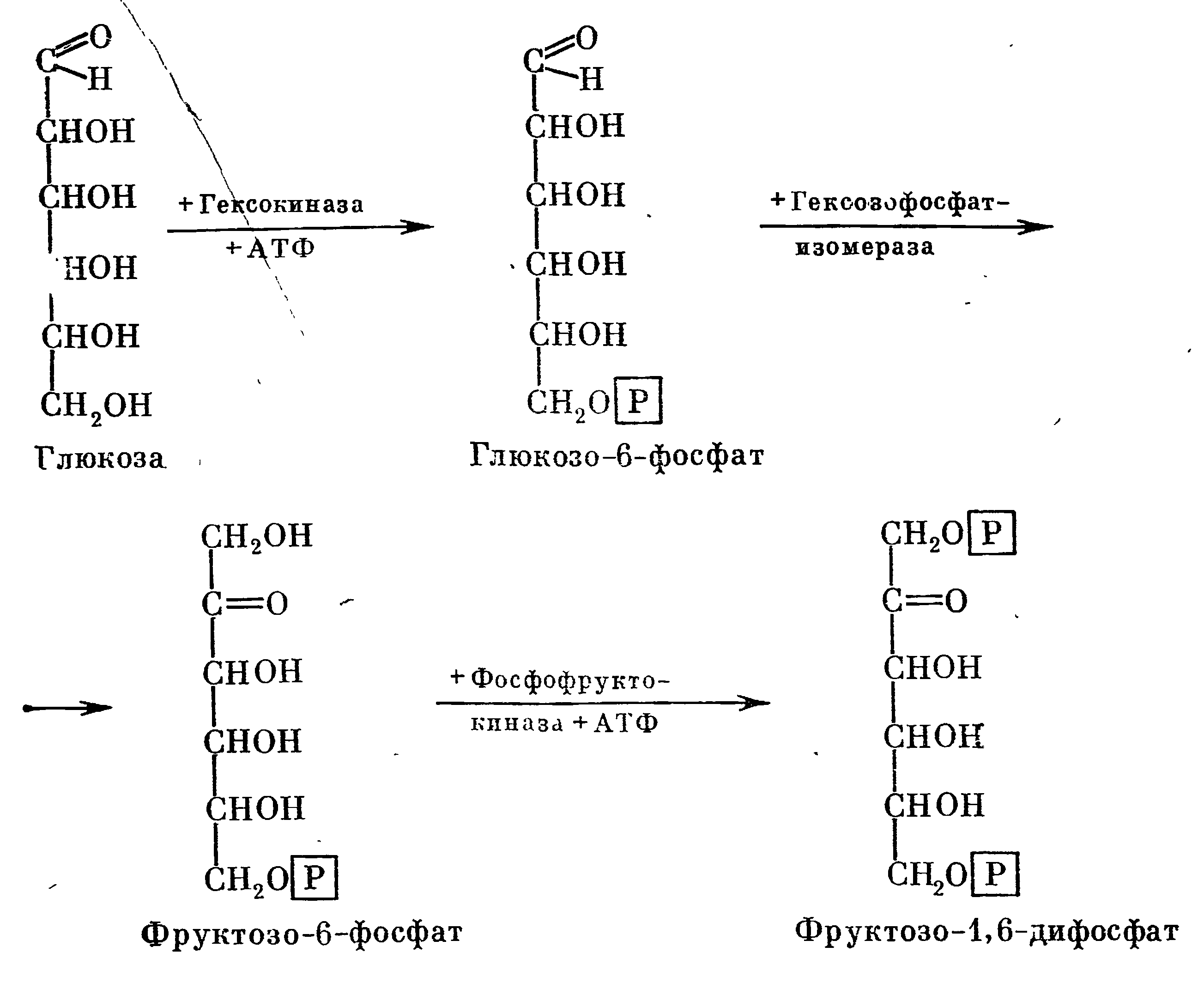
Образованием фруктозо-1,6-дифосфата заканчивается подготовительный этап, который заключается в превращении глюкозы в лабильную форму, способную к дальнейшему превращению.
А. Н. Лебедев первым показал, что фруктозо-1,6-дифосфат расщепляется на глицериновый альдегид и диоксиацетон. Позднее это было подтверждено Г. Эмбденом и О. Мейергофом. Под действием фермента альдолазы фруктозо-1,6-дифосфат распадается на 3-фосфоглицериновый альдегид и фосфодиоксиацетон:
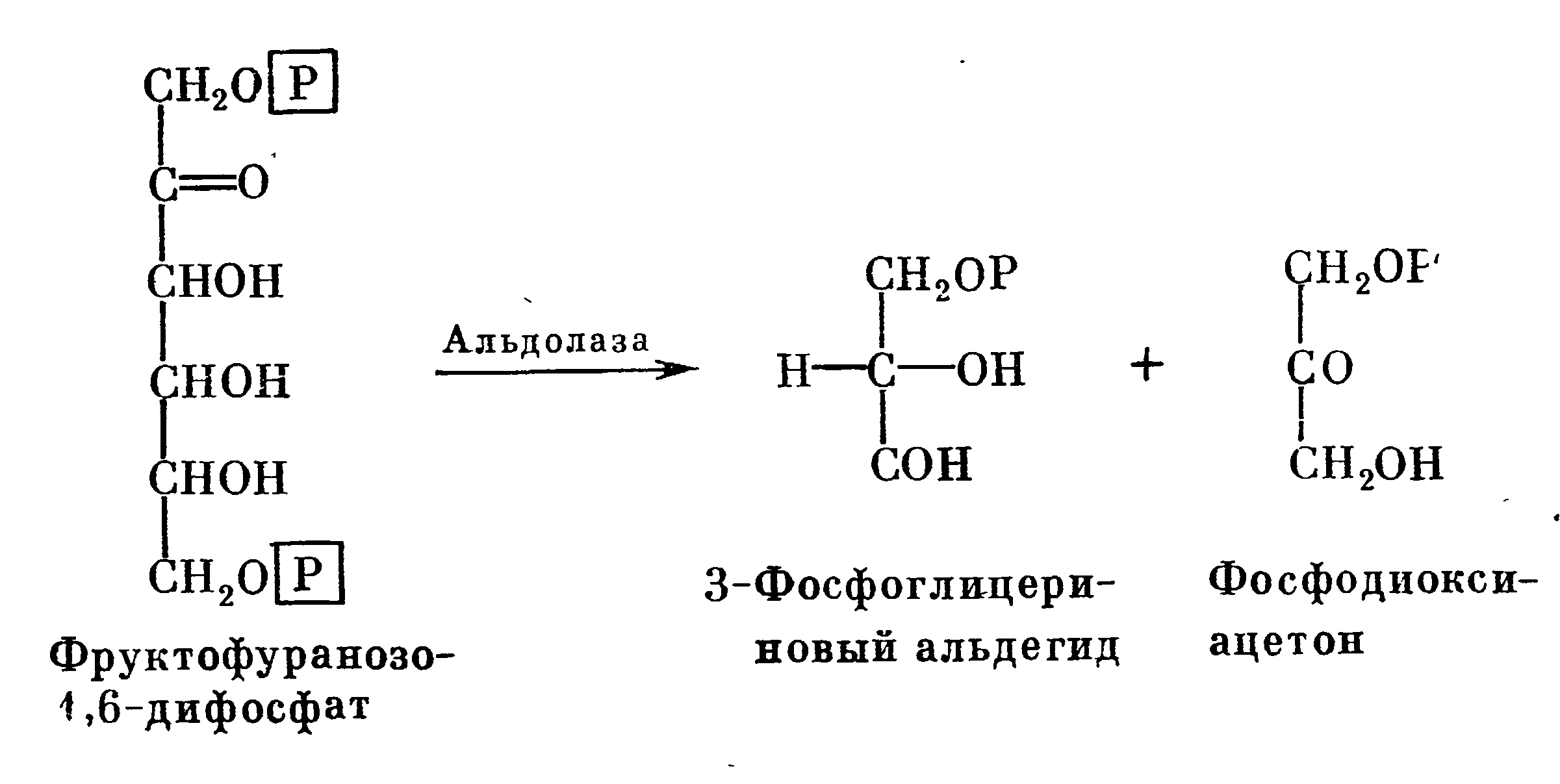
Эти два триозофосфата способны к взаимному превращению под действием фермента триозофосфатизомеразы.
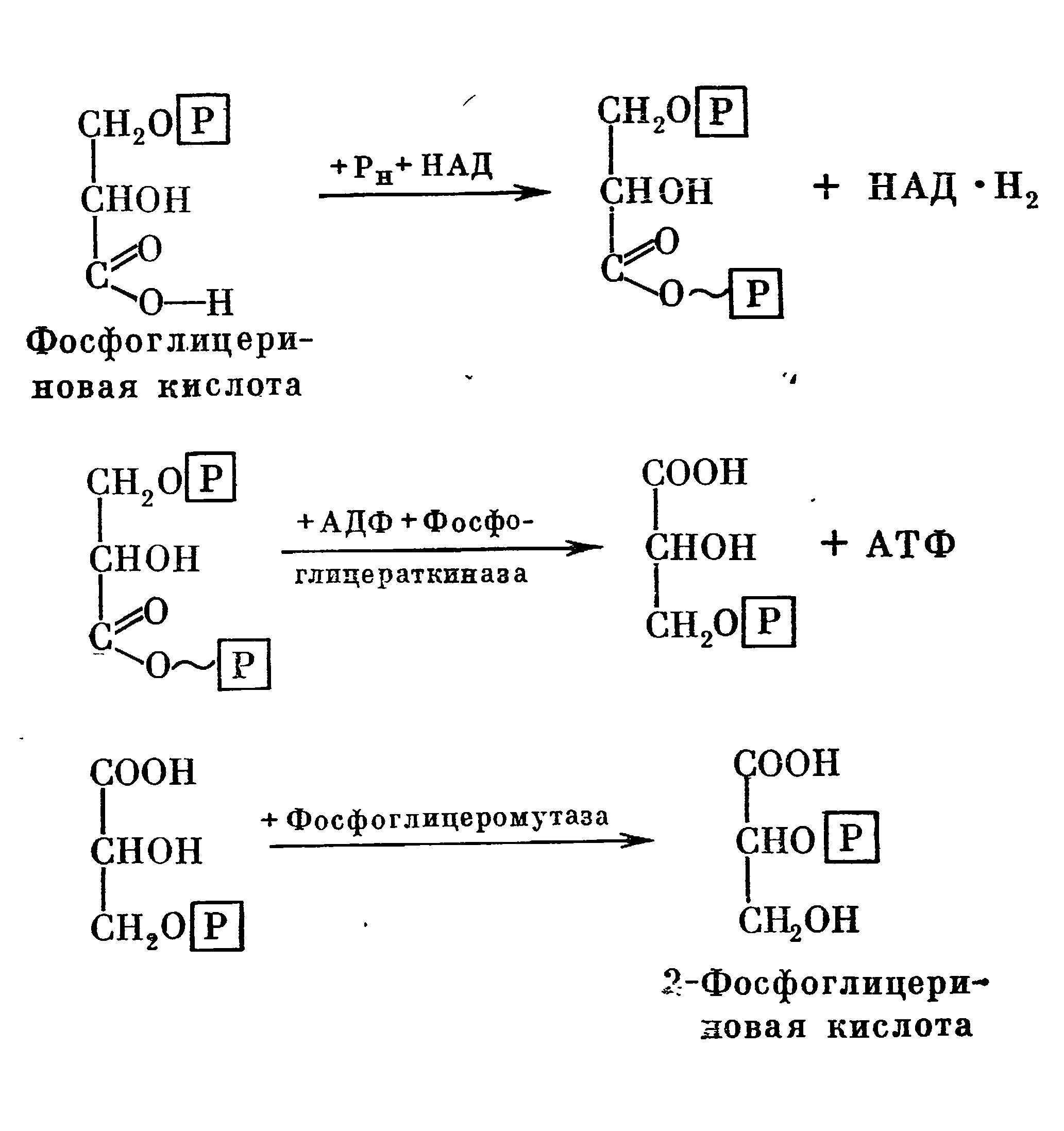
В дальнейшем 3-фосфороглицериновый альдегид присоединяет еще один остаток фосфорной кислоты, за счет неорганического фосфата и образует 1,3-дифосфоглицериновый альдегид, который в присутствии (НАД) окисляется в 1,3-дифосфоглйцериновую кислоту. В этой реакции принимает участие фермент триозофосфатдегидрогеназа.
Образованная 1,3-дифосфоглицериновая кислота под действием фермента фосфоглицерокиназы отщепляет один остаток фосфорной кислоты и передается на АДФ с образованием АТФ и 3-фосфоглицериновой кислоты. Последняя в присутствии фермента фосфоглицеромутазы изомеризуется в 2-фосфоглицериновую кислоту.
Эти реакции можно представить в следующем виде:
Под действием фермента фосфопируватдегидрогеназы 2-фосфоглицериновая кислота превращается в фосфоэнолпировиноградную кислоту, а затем в присутствии фермента фосфотрансферазы фосфоэнолпировиноградная кислота переходит в энолпировиноградную кислоту.
Эти реакции можно изобразить в следующем виде:

Реакция образования пировнноградной кислоты является единственным необратимым процессом во всей цепи биохимических превращений.
Образовавшаяся в процессе брожения пировиноградная кислота может подвергаться различным превращениям в зависимости от наличия анаэробных и аэробных условий, а также от присутствия различных ферментативных систем. При алкогольном брожении пировиноградная кислота действием фермента карбоксилазы декарбоксилируется с образованием уксусного альдегида и углекислого газа:
![]()
И, наконец, последним этапом алкогольного брожения является восстановление уксусного альдегида в этанол.
В этом процессе участвует фермент алкогольдегидрогеназа, который переносит водород с восстановленного НАД-Н2 на уксусный альдегид и образуется этанол.

Восстановление НАД в НАД-Н2 происходит ранее при окислении фосфоглицеринового альдегида в фосфоглицериновую кислоту.
При молочнокислом брожении пировиноградная кислота не декарбоксилируется из-за отсутствия декарбоксилазы у молочнокислых бактерий.
В этом случае она в присутствии лактикодегидрогеназы и НАД-Н2 восстанавливается в молочную кислоту.

Так образуется молочная кислота при молочнокислом брожении и при гликолизе в мышцах животного.
В аэробных условиях пировиноградная кислота участвует во многих синтетических процессах; она превращается в ацетил-КоА, который включается в цикл ди- и трикарбоновых кислот, в синтез жирных кислот и аминокислот.
Таким образом, пировиноградная кислота занимает центральное место в системе реакций, происходящих при анаэробной и аэробной диссимиляции углеводов — в процессе брожения и дыхания.
Энергетический баланс при алкогольном брожении следующий: на каждые 2 моля усвоенного неорганического фосфата образуется 2 моля АТФ при разложении 1 моля сахара.
Глюкоза + 2Ф + 2АДФ→ 2этанола + 2АТФ + СО2 + 2Н2O.
При этом выделяется 117,6 кДж/моль энергии, за счет которой и происходит жизнедеятельность дрожжей.
Дрожжи очень экономно используют энергию при сбраживании глюкозы. В продуктах ее распада остаётся большая часть свободной энергии, которая может быть использована в других биологических процессах при дальнейшем расщеплении этих веществ.
При дыхании глюкоза разлагается до воды и углекислоты по уравнению ![]()
Большую роль сыграли работы С. П. Костычева, посвященные выявлению зависимости между брожением (анаэробным дыханием) и аэробным дыханием. Эта зависимость выражена следующей схемой:
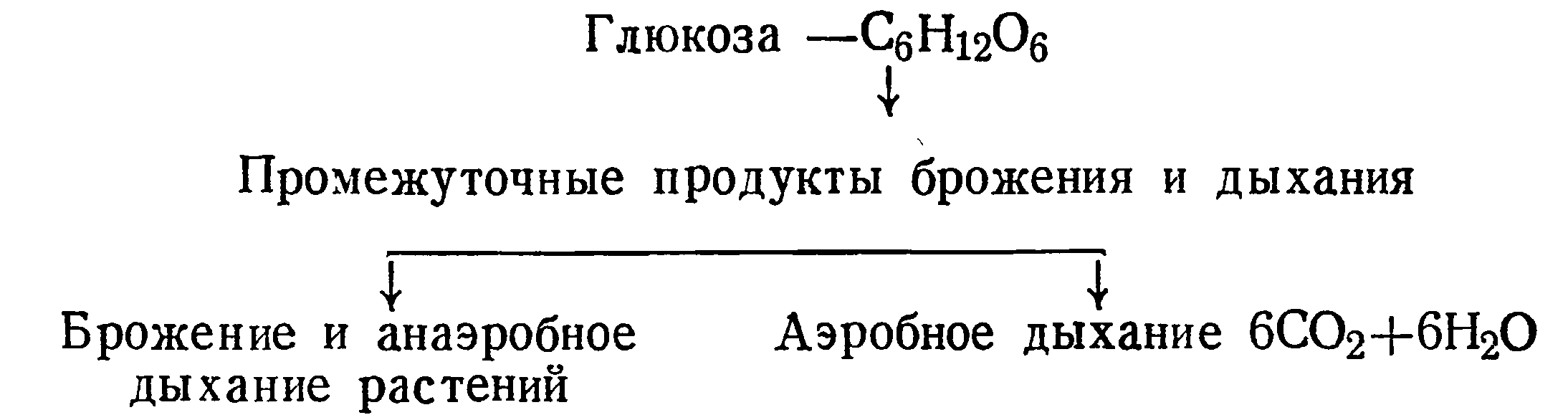
О связи между брожением или анаэробным дыханием и аэробным дыханием свидетельствует то, что в растениях найдены те же промежуточные продукты, которые образуются дрожжами при спиртовом брожении. Это подтверждается тем, что в винограде были найдены этанол, высшие спирты, сложные эфиры, кислоты (пировиноградная кислота) и альдегиды, в том числе и уксусный альдегид. Пировиноградная кислота и уксусный альдегид являются важными продуктами спиртового брожения.
Спиртовое брожение можно регулировать в сторону образования глицерина, как это было показано еще в начале XX в. К. Нейбергом при введении в бродящую жидкость бисульфита натрия, который связывает ацетальдегид (поэтому последний не может служить акцептором водорода). Место уксусного альдегида занимает 3-фосфоглицериновый альдегид, который получает водород от НАД-Н2, образуя глицеринофосфорную кислоту. В результате действия энзима фосфотрансферазы глицеринофосфорная кислота гидролизуется с образованием глицерина и фосфорной кислоты, поэтому вместо этанола в среде накапливается глицерин, уксусный альдегид и углекислота.
Этим и объясняется, что при брожении сусла в присутствии сернистой кислоты образуется значительное количество глицерина и уксусного альдегида.
Активность энзимов дрожжей при спиртовом брожении виноградного сусла меняется [134]. Активность глюкозидазы возрастает с первого дня брожения до пятого, затем падает. Гексокиназа проявляет максимальную активность на третий день брожения. Фосфофруктокиназа и фосфоглюкозоизомераза обнаруживают максимум активности на пятый день, затем их активность падает.
Альдолаза, глицеринальдегид-3-фосфатдегидрогеназа, фосфоглицераткиназа и фосфоглицератмутаза имеют максимальную активность на четвертый день, затем активность их уменьшается, а потом вновь увеличивается.
Кокарбоксилаза (тиаминдифосфат) представляет собой фермент для декарбоксимерования пировнноградной кислоты в ацетальдегид. Она усиливает скорость алкогольного брожения.
Энолаза, пируватдегидрогеназа и алкогольдегидрогеназа достигают максимума активности на четвертый и пятый день после начала брожения. Глюкозо-6-фосфатдегидрогеназа, 6-фосфоглюкон атдегидрогеназа и малатдегидрогеназа проявляют максимум активности между пятым и шестым днями брожения. Активность лактатдегидрогеназы и изоцитратдегидрогеназы уменьшается на четвертый день, а затем начинает возрастать [134]. В этой работе описана методика определения указанных ферментов.
Алкогольдегидрогеназа непосредственно участвует в образовании этанола. Она была выделена из дрожжей в кристаллическом виде; молекулярная масса ее 150 000D, содержит четыре моля НАД, четыре атома цинка и приблизительно 36 свободных SH- групп. Алкогольдегидрогеназа ингибируется йодуксусной кислотой.
Одни авторы считают, что дрожжевые клетки содержат три алкогольдегидрогеназы: одна локализована в митохондриях, две другие — в цитоплазме. Другие полагают, что в дрожжах имеются
две алкогольдегидрогеназы: одна находится в митохондриях, другая — в цитоплазме [176].
Энзиматическое окисление этанола в уксусный альдегид осуществляется митохондриальной алкогольдегидрогеназой. Наоборот, цитоплазматическая алкогольдегидрогеназа в присутствии НАД-Н2 способна восстанавливать уксусный альдегид в этанол. При аэробном окислении этанола дрожжами выделяются ионы водорода (Н+), это сопровождается поглощением из среды эквивалентного количества ионов калия (К+). При этом из этанола образуются эквивалентные количества уксусной кислоты и уксусного альдегида. Установлено, что феназинметасульфат ингибирует поглощение калия (К+) при окислении этанола.
Еще в самом начале XX в. Г. Виланд показал, что дрожжи Sacch. cerevisiae окисляют этанол в аэробных условиях в уксусный альдегид, а затем в уксусную кислоту. При интенсивной аэрации дрожжи окисляют уксусную кислоту до углекислоты и воды, при этом расход спирта на 20% больше, чем образующихся конечных продуктов. Объясняется это тем, что значительные количества этанола используются дрожжами для синтеза биомассы углеводов и липидов. Дрожжи способны дегидрировать и другие высшие спирты. Это свидетельствует о том, что в вине присутствуют высшие альдегиды алифатического ряда.