Глава 13
МЕТАБОЛИЗМ ОРГАНИЧЕСКИХ КИСЛОТ ДРОЖЖАМИ ПРИ БРОЖЕНИИ
Органические кислоты являются продуктами диссимиляции углеводов, аминокислот и жиров микроорганизмами. Существуют два пути превращения углеводов: аэробнопентозофосфатный и анаэробно-гликолитический.
В результате углеводного и азотистого обмена у дрожжей в процессе алкогольного брожения образуется целый ряд веществ, играющих важную роль в формировании вина. Среди этих веществ на одном из первых мест стоят органические кислоты, которые играют определенную физиологическую роль, в значительной степени определяют внутренние условия среды (pH) в дрожжевой клетке и регулируют внутреннюю интенсивность и направленность биохимических реакций.
В зависимости от соотношения свободных и связанных в соли кислот находится актуальная кислотность и концентрация водородных ионов.
Биосинтез органических кислот
Основные представления о путях превращений органических кислот в растениях и микроорганизмах изложены в работах
С. П. Костычева, В. С. Бутуевича, М. В. Федорова, Т. Кшонца, К. Бергауэра, X. Кребса и В. Джонсона, X. Корберга и X. Кребса, Д. Девиса и др.
С. П. Костычев [53] на заре развития биохимии считал, что образование лимонной кислоты немыслимо без синтетических реакций. Впоследствии это было подтверждено советскими и зарубежными исследователями.
Позднее было доказано, что синтез лимонной кислоты протекает по циклу Кребса посредством конденсации щавелевоуксусной кислоты с ацетил-КоА. Был выделен фермент, который катализировал конденсацию щавелевоуксусной кислоты с ацетил- КоА, поэтому этот фермент был назван конденсирующим энзимом.
Образование лимонной кислоты из глюкозы — сложный биохимический процесс. Вначале из глюкозы образуется пировиноградная кислота, но этот процесс чрезвычайно сложен.
.Изучение анаэробного распада углеводов привело к установлению следующих промежуточных продуктов: углеводы + фосфорная кислота + гексозофосфат → триозофосфат + фосфоглицериновая кислота → фосфопировиноградная кислота. Дальнейшее превращение пировнноградной кислоты в зависимости от наличия ферментативных систем у дрожжей протекает различно.
Известно, что пировиноградная кислота является исходным продуктом для синтеза целого ряда важнейших веществ, определяющих направление обмена в живой клетке.
Дрожжи способны декарбоксилировать пировиноградную кислоту в уксусный альдегид, они также способны непосредственно превращать пировиноградную кислоту в уксусную посредством окислительного декарбоксилирования.
При спиртовом брожении пировиноградная кислота через уксусный альдегид образует этанол, а при гликолизе под действием лактикодегидрогеназы и НАД-Н2 она превращается в молочную. Пировиноградная кислота участвует в реакциях переаминирования, она легко аминируется в аланин.
Пировиноградная кислота участвует в синтезе кислот по циклу Кребса. Но прежде, чем она будет вовлечена в циклы ди- и трикарбоксильных кислот, она должна, с одной стороны, карбоксилироваться в щавелевоуксусную кислоту, а с другой — в присутствии коэнзима А превратиться в ацетил-КоА. Эти две реакции можно представить в следующем виде:
![]()
Коэнзим А является переносчиком ацетильной группы, которая присоединяется к нему через эфирную микроэргическую связь. Поэтому в микроорганизмах широко распространены ацетилфосфаты, которые принимают участие в биохимических реакциях, связанных с выделением энергии. Роль коэнзима А в биологических окислениях и синтезах стала более интенсивно исследоваться после препаративного приготовления его, осуществленного Ф. Линеном и Е. Рейхертом. Была установлена взаимная связь между изменением содержания коэнзима А в дрожжевых клетках и скоростью окисления уксусной кислоты. Доказано, что при недостатке коэнзима А дрожжи слабо окисляют уксусную кислоту.
Ацетил-КоА образуется не только из пировиноградной кислоты, но и из уксусной кислоты и уксусного альдегида. Источником образования щавелевоуксусной кислоты являются яблочная и аспарагиновая кислоты, а также путь карбоксилирования пировиноградной кислоты.
Дрожжи обладают всеми ферментативными системами для образования щавелевоуксусной кислоты из указанных соединений, они содержат также коэнзим А и способны образовывать ацетил- КоА из пировиноградной и уксусной кислот.
С помощью изотопного углерода в виде NaHC14O3 нам в 1959 г. удалось доказать, что винные дрожжи способны карбоксилировать пировиноградную кислоту с образованием щавелевоуксусной кислоты. Последняя реагирует с ацетил-КоА и образует лимонную кислоту по схеме
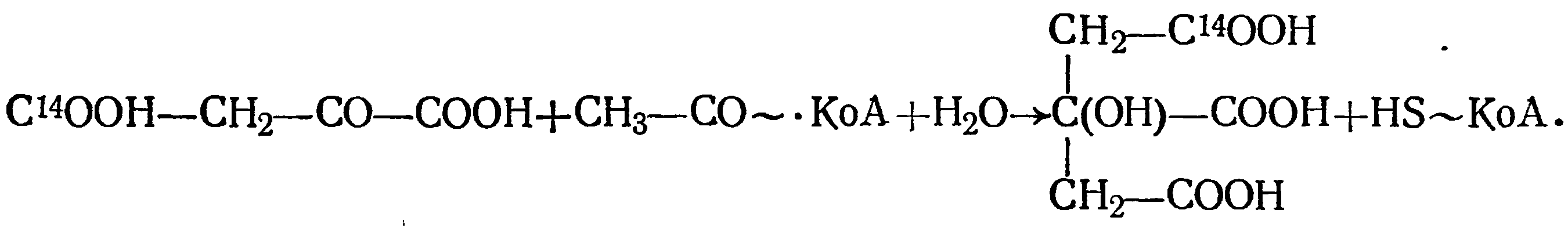
Методом бумажной хроматографии из культуральной жидкости дрожжей были выделены радиоактивная лимонная кислота, молочная, яблочная и янтарная.
Винные дрожжи способны превращать лимонную кислоту по циклу трикарбоксильных кислот и образовывать янтарную и глиоксалевую кислоты, последняя конденсируется с ацетил-КоА и образует яблочную кислоту по глиоксалатному циклу.
Кроме того, винные дрожжи также способны синтезировать лимонную кислоту из глютаминовой кислоты и аланина путем дезаминирования их в соответствующие кетокислоты и в дальнейшем превращать их по циклу Кребса.
Во-первых, винные дрожжи способны карбоксилировать пиро- виноградную кислоту в щавелевоуксусную. Они также способны превращать пировиноградную кислоту в ацетил-КоА. Последний конденсируется с щавелевоуксусной кислотой и образует лимонную кислоту.
Во-вторых, дрожжи путем окислительного декарбоксилирования превращают пировиноградную кислоту в уксусную. Последнюю они способны конденсировать в янтарную кислоту через ацетил-КоА, янтарную кислоту они превращают в щавелевоуксусную через фумаровую и яблочную. Щавелевоуксусная кислота конденсируется с ацетил-КоА и образуется лимонная кислота.
В работе С. В. Дурмишидзе (1965) было показано, что меченая уксусная кислота превращается в янтарную, фумаровую и гликолевую кислоты. По данным автора процесс ассимиляции и превращения уксусной кислоты протекает на всех стадиях алкогольного брожения. Известно также, что уксусная кислота в виде ацетил-КоА через цикл ди- и трикарбоновых кислот окисляется до СO2 и воды.
Генезис уксусной кислоты при алкогольном брожении долгое время оставался неясным и только благодаря работам С. П. Костычева стало известно, что дрожжи превращают уксусный альдегид и в этанол и уксусную кислоту по следующей схеме:
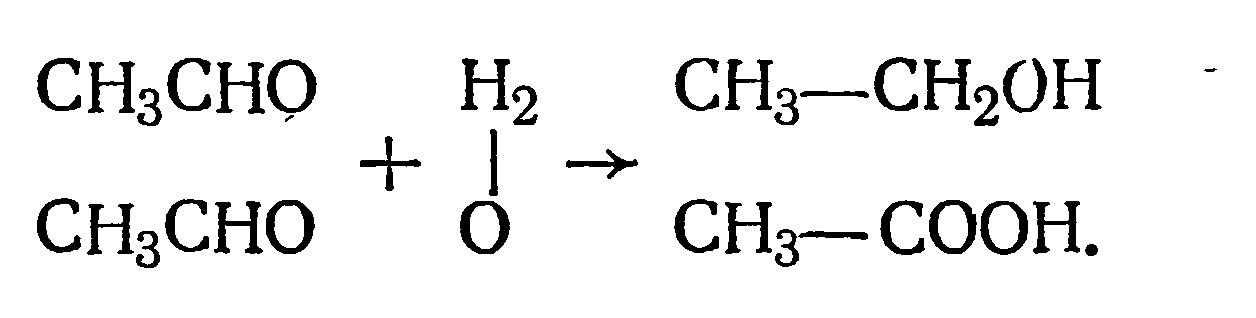
По современным представлениям эта реакция протекает в два этапа: на первом уксусный альдегид присоединяет кислород воды и окисляется до уксусной кислоты, при этом водород воды восстанавливает НАД, который участвует в этой реакции: СН3СНO+Н2O+НАД→СН3СOOН+НАД.H2.
Образовавшийся НАД-Н2 восстанавливает вторую молекулу уксусного альдегида в спирт: СН3СНО+НАД-Н2→СН3—СН2ОН.
Исследования Е. Пейно показали, что в начале брожения количество летучих кислот, в частности уксусной, заметно возрастает, затем процесс замедляется, количество летучих кислот стабилизируется и в конце брожения начинает убывать. Этот автор считает, что в период интенсивного размножения дрожжей их восстановительная способность высока, поэтому и происходит восстановление уксусной кислоты в этанол.
Известно, что при усилении восстановительных процессов происходит максимальное накопление этанола. Однако трудно понять непосредственное восстановление уксусной кислоты в этиловый спирт, как это понимает Е. Пейно.
Е. Пейно использовал способность дрожжей восстанавливать уксусную кислоту в этанол в анаэробных условиях для исправления вин с высоким содержанием уксусной кислоты/ как это было сделано Η. Ф. Саенко для лечения больных вин хересными дрожжами.
Образование уксусной кислоты в процессе спиртового брожения сусла зависит от pH среды и от температуры брожения. При брожении в менее кислой среде (pH 6) вначале образуется значительное количество уксусной кислоты, затем скорость образования уменьшается. При pH 7,5 она образуется равномерно в течение всего периода, а при pH 2,5 она в вине не накапливается.
Е. Пейно исследовал 32 расы дрожжей на способность образовывать летучие кислоты в присутствии кислорода и без него. Среднее количество образующихся при брожении в аэробных условиях летучих кислот составляло 0,52 г/л, а в анаэробных условиях — 0,69 г/л.
Г. Г. Валуйко также наблюдал, что в присутствии кислорода летучих кислот образуется меньше. Известно, что аэробиоз подавляет образование летучих кислот. Объясняется это тем, что при сбраживании сусла в аэробных условиях уксусная кислота окисляется в ацетил-КоА через цикл Кребса.
В основном уксусная кислота образуется не только в процессе брожения, но и при ферментативном окислении спирта. Это происходит при выдержке виноматериалов с дрожжевым осадком. Процесс усиливается при аэробных условиях, т. е. если бочки не доливать вином до шпунта.
Уксусная кислота образуется также при β-окислении высокомолекулярных жирных кислот. При этом окисление происходит у того углеродного атома, который находится в β-положении по отношению к карбоксильной группе:![]()
В результате разрыва цепочки жирной кислоты между а- и β-углеродными атомами образуется уксусная кислота; новая высокомолекулярная кислота будет содержать на 2 атома углерода меньше, чем исходная жирная кислота.
Известно, что при более высоком pH среды образуется меньше 2,3-бутиленгликоля и ацетоина и значительно больше — глицерина, летучих кислот и янтарной кислоты. Процесс образования янтарной кислоты очень сложный. Превращения улусной кислоты начинаются с образованием ацетил-КоА. Конденсируются не две молекулы уксусной кислоты, а активные ацетильные группы в виде ацетил-КоА, которые и образуют янтарную кислоту.![]()
Как показали исследования В. З. Гваладзе, при брожении сусла в анаэробных условиях количество янтарной кислоты бывает незначительно, а в аэробных условиях увеличивается.
Янтарная кислота образуется также при расщеплении изолимонной кислоты при действии изоцитратазы по схеме:

Глиоксалевая кислота, согласно глиоксалатному циклу, вступает в реакцию с ацетил-КоА и образует яблочную кислоту по схеме
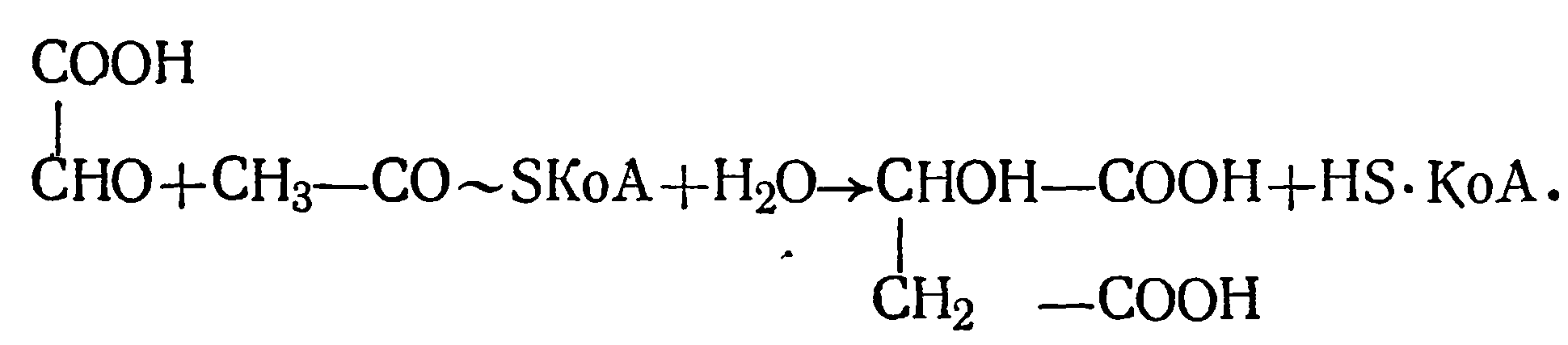
Эти три кислоты были найдены в винограде и вине.
В вине была обнаружена α-метиляблочная кислота, или лимонно-яблочная. Она образуется в результате конденсации пировнноградной кислоты с ацетил-КоА (Ж. Карл, 1969):
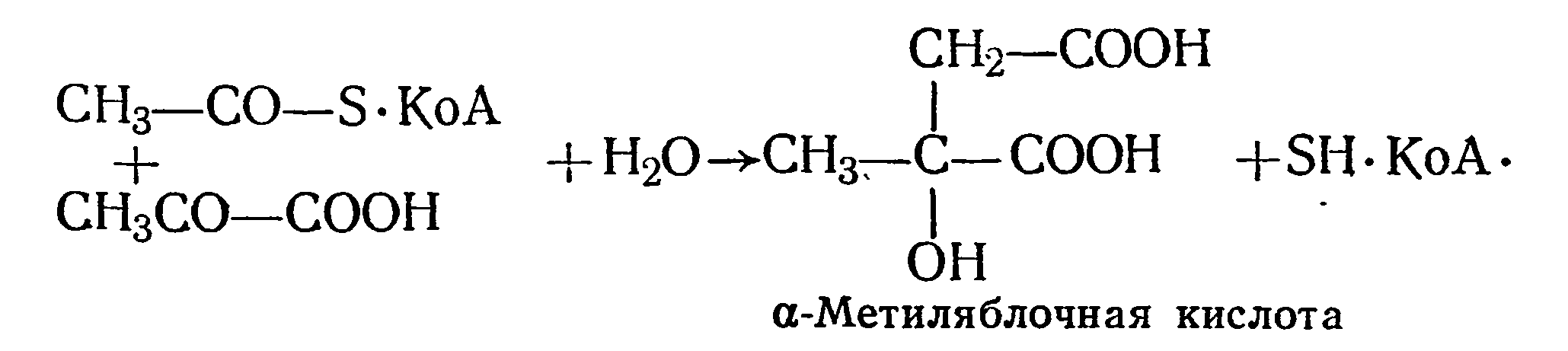
Существует и другой путь образования α-метиляблочной кислоты — это конденсация глиоксалевой кислоты с пропионилом- КоА с образованием α-оксиглутаровой кислоты. Последняя в результате внутримолекулярной перегруппировки превращается в метил яблочную кислоту по следующей схеме:
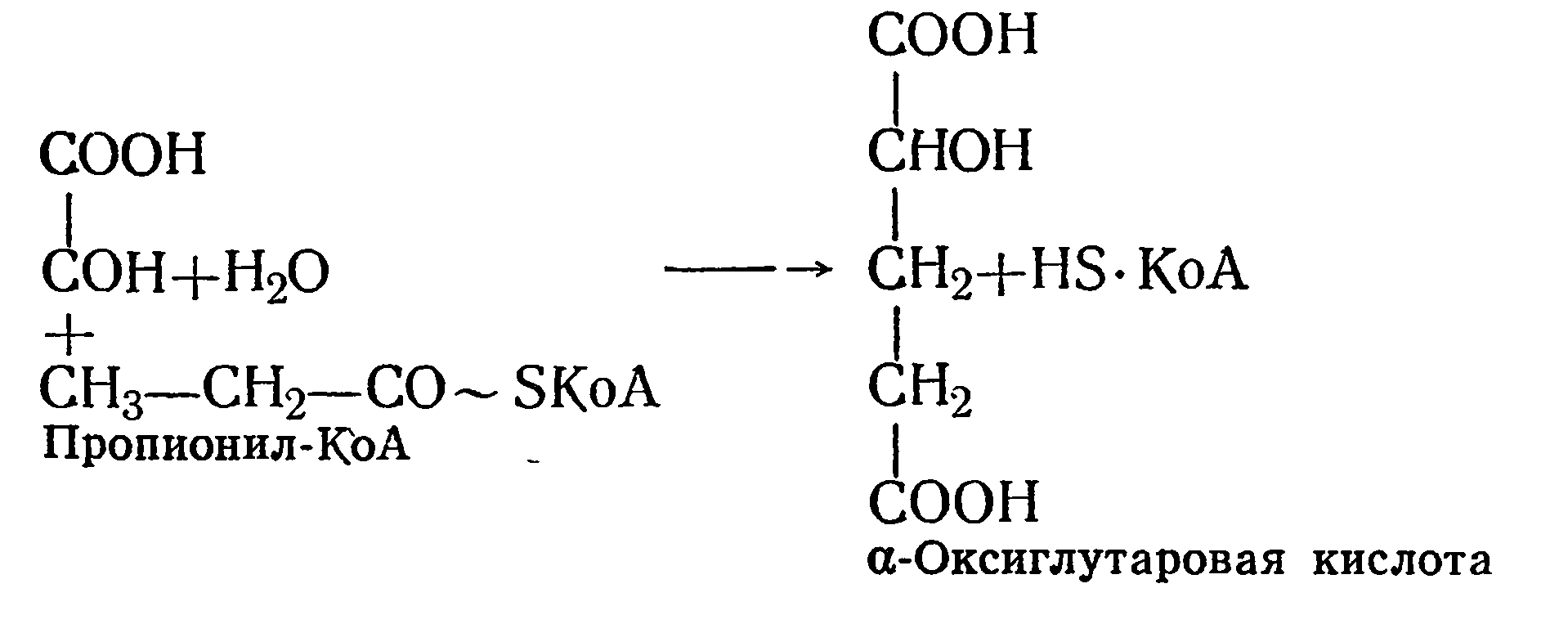
α-Оксиглутаровая кислота легко распадается на а-метиляблочную кислоту, а последняя — на пировиноградную и уксусную кислоты:
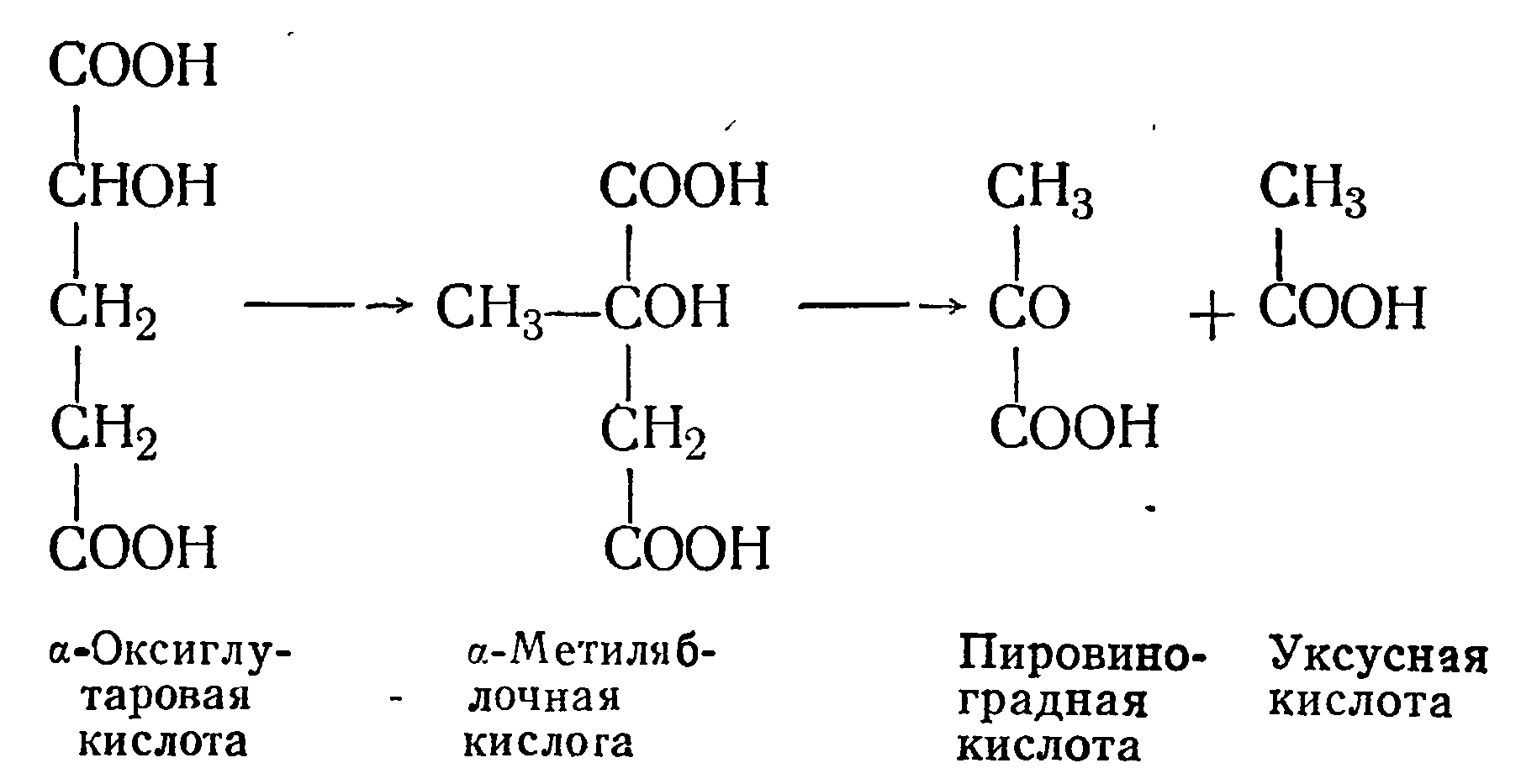
α-Метиляблочная кислота образуется также при декарбоксилировании лимонной кислоты винными дрожжами по схеме
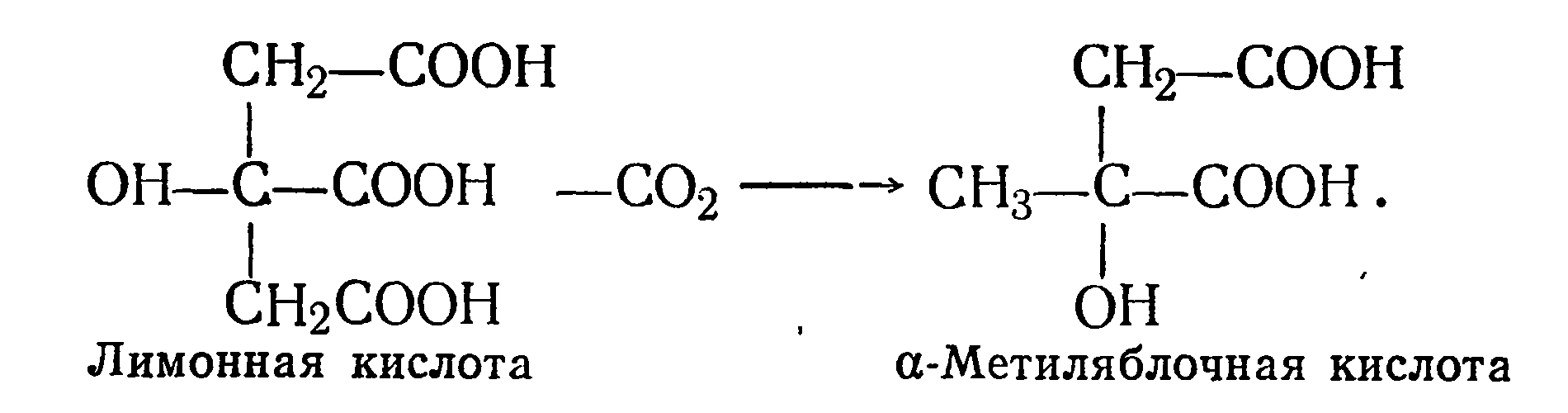
Исследования показали, что α-метиляблочная кислота не содержится в винограде. Она образуется при алкогольном брожении из сахара в количестве от 70 до 180 мг/л. Концентрация ее прямо пропорциональна содержанию спирта и зависит от условий брожения, в частности от метаболитической активности дрожжей.
Количество α-метиляблочной кислоты зависит также и от pH среды: при pH 3,9 ее накапливается до 80, а при pH 3,2 до 282 мг/л. Количество α-метиляблочной кислоты в белых винах
колеблется от 65 до 147 мг/л и в красных — от 91 до 187 мг/л. Содержание ее значительно увеличивается при выдержке вина.
В вине найдены 2,3-диоксивалериановая, диметилглицериновая, или 2-метил-2,3-диоксимасляная кислоты. Последняя образуется в результате конденсации пировнноградной кислоты с уксусным альдегидом:
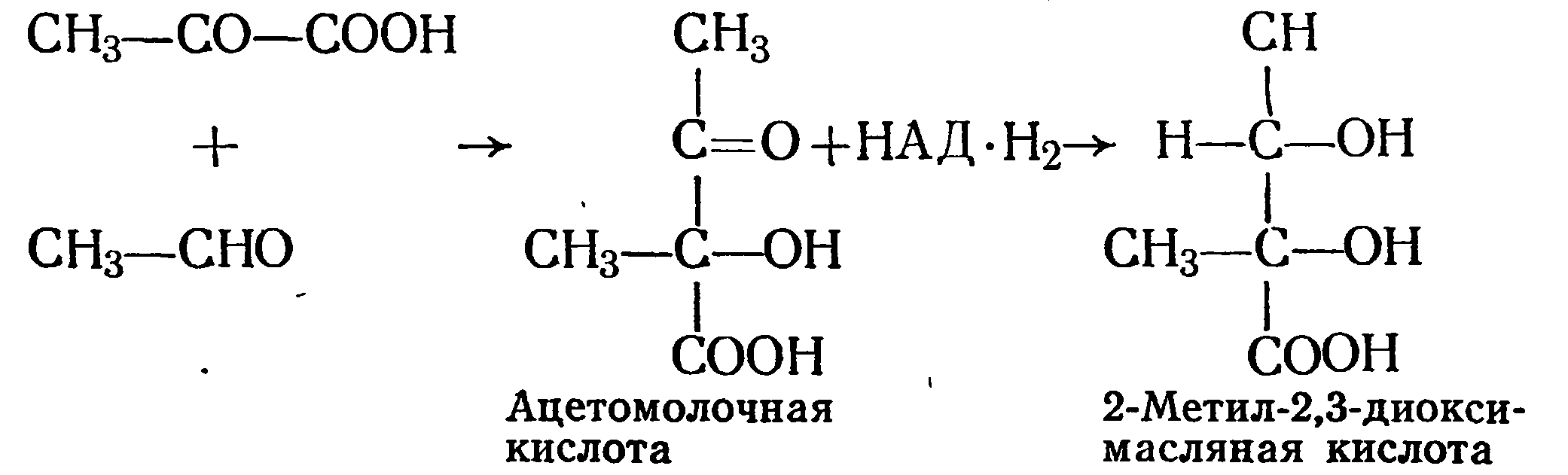
Образованная ацетомолочная кислота восстанавливается в присутствии НАД-Н2 в 2-метил-2,3-диоксимасляную кислоту.
Установлено, что содержание α-метиляблочной, диметилглицериновой и 2-метил-2,3-диоксимасляной кислот зависит от расы дрожжей и их метаболитической активности. Накопление двух последних кислот в винах зависит также от условий проведения брожения. Так при анаэробиозе количество их уменьшается.
В винах были найдены гликолевая, глиоксалевая, фумаровая, пировиноградная, щавелевоуксусная, глюконовая, галактуроновая кислоты. Что касается глюкуроновой кислоты, то, по мнению одних авторов, в обычных винах ее нет. Она встречается в винах, приготовленных из винограда, пораженного грибом Ботритис цинерея, при этом количество ее достигает 1 г/л и больше. Она образуется в результате окисления глюкозы ферментами этого гриба. По мнению других, глюкуроновая кислота встречается в бордоских сухих винах в количестве 100—200 мг/л.
В выдержанных винах содержится диоксифумаровая кислота. В аэробных условиях она быстро распадается на мезоксалевую, глиоксалевую, гликолевую и щавелевую кислоты.
Согласно данным П. Риберо-Гайона, содержание ароматических кислот (n-оксикоричной, n-оксибензойной и салициловой) в красных винах составляет от 50 до 100, а в белых — до 5 мг/л [154].
По данным этого же автора, в винах встречаются следующие фенольные кислоты: шикимовая, хинная, ферулевая, галловая, кофейная и протокатеховая. Биосинтез фенольных соединений был показан ранее.
В наибольшем количестве в винах встречаются жирные кислоты. Они находятся в свободном состоянии, в виде сложных эфиров, а также в виде солей. Низшие жирные кислоты (от С1 до С9) при обычных условиях являются жидкими. Обычно, начиная с масляной кислоты до С9, они представляют собой маслянистые жидкости и легко растворяются в воде в определенных концентрациях. Высшие жирные кислоты (от С10 и выше) являются твердыми веществами.
Дрожжевая клетка содержит все ферменты, катализирующие синтез и распад жирных кислот. Основными жирными кислотами вина являются уксусная, н-капроновая1, н-каприловая, н-каприновая и дециловая. В меньшем количестве встречаются муравьиная, пропионовая, изомасляная, α-метилмасляная, 2-метилдиоксимасляная, изовалериановая, 2-оксикапроновая и н-пеларгоновая.
Из высокомолекулярных кислот в винах содержатся лауриновая, пальмитиновая, стеариновая, олеиновая, линолевая и др.