Вирус относится к группе бороздчатости древесины яблони (Apple stem grooving virus groop ).
Криптограмма. */*:*/*:E/E:S/*
Вирусные частицы нитевидной формы, размером 600—700 X Х12 nm, коэффициент седиментации 120 S. Температура инактивации 60—67°С, разведение 10-2—10-3, в соке сохраняет инфекционность два—четыре дня, выдерживает промораживание, умеренный или плохой антиген, переносчик не известен.
Поражаемые растения.
Яблоня (Malus sylvestris, М. hupehensis, M. sieboldii, M. sieboldii var. arborescens), груша (Pyrus communis), айва (Cydonia oblonga), Pyronis veitchii (латентно) и 16 видов травянистых растений.Вирус бороздчатости вызывает на древесине индикатора Virginia crab широкие плоские борозды, ямчатость, некротическую линию или отдельные бурые некротические пятна в зоне спайки индикатора с подвоем (рис. 22,а, г). Часто наблюдаются также расположение привоя под более тупым углом к подвою и образование утолщения выше спайки. Побеги индикатора V. crab, зараженные вирусом бороздчатости, часто обламываются, а позже начинают чахнуть и отмирать. Рост у зараженных растений слабый, листья хлоротичные, рано краснеющие, плоды созревают преждевременно и ярко окрашены (рис. 22,б). Отсюда описание этого вируса под названиями «отмирание V. crab» и «атипичная некротическая ямчатость древесины» (Welsh, Keane, 1959 а; Welsh, Nyland, 1965).
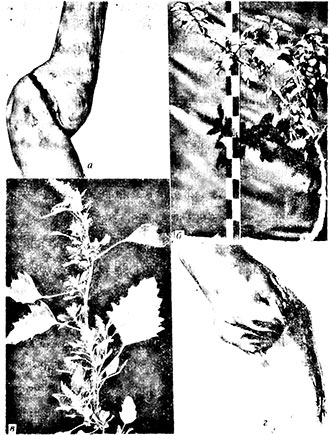
Рис. 22. Вирус бороздчатости древесины яблони:
а, г — симптомы бороздчатости и бурой линии в зоне спайки индикатора V. crab с подвоем; б — атрофия кроны индикатора V. crab; в — кустистость и зеленые эпинастии Ch. quinoa, зараженного вирусом бороздчатости древесины
Распространение и история изучения.
Описан в большинстве стран Европы и Северной Америки, не установлен в Австрии, Югославии, Франции, Турции, Греции и Мексике. В Советском Союзе вирус бороздчатости древесины выявлен в Молдавии, на Украине, в (Белоруссии, Латвии и Центральной черноземной зоне.
Вирус впервые описали О. A. Sequeira (1965) под названием «Е-36 вирус» и R. Lister под названием «Изолят С-431» (Listei Nadakavukaren, Bancroft, 1965), а позднее Η. E. Waterworth, R. M. Gilmer (1969), под названием вируса темно-зеленых эпинастий. О. A. Sequeira (1967) первый выполнил пассаж выделенного им изолята Е-36 с травянистых на древесные индикаторы и наблюдал на зараженном таким образом V. crab симптомы бороздчатости ствола, ненормальности и некрозы в месте прививки.
Дальнейшее изучение изолятов Е-36 и С-431, а позже вируса темно-зеленых эпинастий показало, что все они родственны и представляют собой образцы одного вируса, который решено было назвать вирусом бороздчатости ствола яблони (Apple stem grooving virus).
Два образца этого вируса перенесены с травянистых на древесные индикаторы и через 30 месяцев после заражения получили симптомы бороздчатости и бурой линии на V. crab. Второй изолят не вызвал симптомов через 42 месяца, поскольку, по мнению этих авторов, был суперлатентным (Gilmer, Mink, 1971).
В отличие от названных авторов F. A. Van der Meer (1976) исследовал 11 чистых изолятов вируса бороздчатости, среди которых были как очень сильные, так и очень слабые штаммы. Однако индикатор Virginia crab в его опытах выявлял все изоляты. В дальнейших исследованиях автор раскрыл причины разногласий в ранних опытах М. Welsh и G. Nyland о взаимосвязи вируса бороздчатости древесины и образования бурой линии в зоне спайки. Как показал F. A. Van der Meer (1982), бурая линия — это реакция сверхчувствительности индикатора V. crab на заражение вирусом бороздчатости древесины. В тех случаях, когда вирус инфицировали на подвой, бурая линия не развивалась, так как вирус не распространялся системно в подвое, а оставался локализованным возле инфекционной почки. Прекрасным доказательством связи вируса бороздчатости древесины с образованием некротической линии в зоне спайки со сверхчувствительным привоем служат эксперименты японского исследователя Н. Yanase (1982). Он вместо V. crab использовал гиперчувствительные виды М. sieboldii и М. sieboldii var. arborescens, заразил их очищенными препаратами вируса и получил прекрасную некротическую реакцию.
В ЮАР получены данные о том, что вирус бороздчатости способен вызывать явление несовместимости и гибель деревьев груши сорта Пакхам Триумф в условиях естественного заражения (Van Siebert, Engelbrecht, 1982), что заставляет по-новому взглянуть на явление несовместимости и отмирание деревьев груши.
В Молдавской ССР вирус бороздчатости описан в 1969 г. Зараженность сортов и подвоев составляет 20%. При этом вирус бороздчатости всегда присутствовал в комплексе с двумя другими латентными вирусами: вирусом хлоротической пятнистости листьев и вирусом ямчатости древесины. Чистые изоляты этого вируса получены после термотерапии, где он часто оставался после инактивации всех других вирусов. Как правило, изоляты вируса бороздчатости древесины удается инактивировать только после двухтрехразовой тепловой обработки.