Общие сведения
Покоящееся состояние растительных органов физиологи (Иванов, 1936; Максимов, 1958; Рубина, 1952) противопоставляют состоянию роста. Отсюда следует, что явление покоя присуще только таким органам и тканям, которые способны к росту, т. е. имеют меристематическое сложение. К таковым у заканчивающих рост побегов относятся эмбриональные ткани почек и камбий.
Как отмечалось выше, почки, находящиеся в пазухе пасынка, прорастают, как правило, весной после зимовки, поэтому их называют "зимующими почками" или "зимующими глазками". Состояние, в котором находятся почки винограда осенью и зимой, называется состоянием покоя. По определению Генкеля и Окниной (1954), покоем у растений называется состояние, при котором почти полностью приостанавливаются процессы роста, снижается интенсивность обмена веществ и содержимое клеток претерпевает глубокие изменения.
Максимов (1958) определяет понятие покоя следующим образом: "В покоящихся органах, говорит Максимов, можно обнаружить все важнейшие проявления жизнедеятельности, кроме одного, именно роста, который не идет, даже если им предоставить вполне благоприятные для роста условий".
Хотя значение пониженной температуры осенне-зимнего периода остается не вполне выясненным для дальнейшего развития зимующих почек, из многих работ различных авторов можно заключить, что если не подвергнуть их воздействию пониженных температур, то они не приобретают способности к прорастанию весной и не будут развиваться дальше (Кондо, 1955). Определяющим моментом, утверждает Кондо (1969), в состоянии покоя зимующих почек в конце вегетационного периода является потребность в холоде. В случае же неудовлетворения ее меристематические клетки не могут пройти соответствующие обменные реакции, подготовляющие переход почек к активному росту, к новой вегетации.
При выдерживании ранней осенью виноградных черенков в холодильнике при температуре близкой к 0°С, органический покой почек длится около полутора месяцев. Растянутость периода органического покоя на виноградниках объясняется тем, что в природных условиях температура сохраняет свое оптимальное для прохождения покоя значение в пределах ограниченного времени — осенью (особенно днем) она долго держится выше необходимого оптимума, а зимой — ниже.
Потапенко и Костина (1950) считают, что для того, чтобы приобрести свойства прорастания, почки должны испытать воздействие пониженной температуры в пределах 8—0°С. Продолжительность воздействия пониженной температуры специфична для отдельных сортов и обуславливается их эколого-географическим происхождением. Magoon и Dix (1943) установили, что распускание почек ускоряется тем больше, чем дольше зимой, до перемещения в теплицу, они оставались в условиях открытого грунта. Вranas с сотр. (1946), сопоставляя метеорологические условия с датой распускания почек кустов, зимовавших в условиях открытого грунта, пришли к выводу, что с продлением периода похолодания уменьшается количество тепла, требуемого на распускание почек. Nigоnd (1957) также отмечает, что температура в интервале от 1 до 8°С оказывает более эффективное воздействие на сокращение органического покоя, чем температура на уровне 0°С, однако воздействие температурой 18 или 25°С оказалось еще более эффективным.
Alleweldt (1960) установил, что воздействие температурой +5 или —5°С в течение 15 дней не сокращает срок прорастания почек и не повышает процент распустившихся глазков. К аналогичным выводам приходят также Antcliff и May (1961). Почки Султанина, по их мнению, не требуют воздействия низкими температурами для приобретения способности к прорастанию.
Ряд исследователей (Потапенко, 1949; Потапенко, Костина, 1950; Тютюник, 1953) считает, что почки начинают усваивать понижение температуры очень рано — еще в конце лета и начале осени.
Следует отметить, что в литературе не выяснен вопрос о параметрах пониженной температуры, необходимой для оказания требуемого физиологического воздействия на прорастание почек весной. В работах Гулисашвили (1954) указано, что продолжительность воздействия пониженной температуры составляет 57 дней, причем в это время не требуется, чтобы температура падала ниже +1°С.
Для прорастания почек, находящихся в фазе т. наз. условного — корреляционного покоя, не обязательно прохождение периода пониженной температуры. Мержаниан (1939) давно сообщил, что прищипка верхушек побегов и удаление пасынков примерно до конца июля вызывает развитие зимующих почек в год их формирования. Негруль (1952) также отмечает, что при удалении всех точек роста можно вызвать развитие зимующих почек в год их формирования.
Многим исследователям впоследствии удалось вызвать развитие зимующих почек в год их формирования (до вступления в органический покой) и наряду с этим доказать, что для их прорастания в фазе условного покоя вовсе не требуется воздействия пониженной температурой. Здесь мы снова возвратимся к исследованиям Стоева и Никова (1956), которые показали, что удаление всех точек роста у сортов Памид, Димят, Болгар, Тамянка, Пино черный и Уньи белый с 15 /VI до 15 /VII вызывает массовое прорастание зимующих почек. В конце июля (в начале вызревания побегов), однако, прорастание почек уменьшилось на 15—30%, 15/VIII процент проросших почек снизился до 20—30, а к середине сентября авторам удалось вызвать прорастание лишь единичных почек.
Таблица 12
Динамика прорастания зимующих почек во время условного покоя
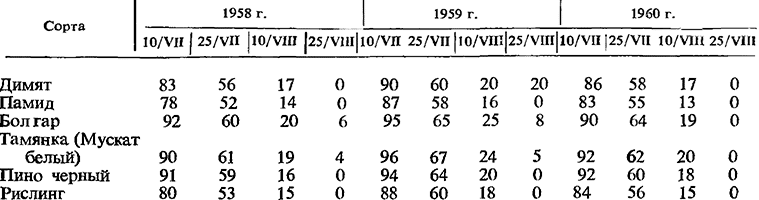
Таблица 13
Прорастание зимующих почек (%) в зависимости от их местоположения по длине побега

Динамика прорастания зимующих почек после 10/VII, по данным Никова (1964), представлена в табл. 12 (прорастание до 10/VII составляло 100%).
Ников (1964) установил также, что потеря способности к прорастанию почек различных ярусов наступает неодновременно. Как правило, низко расположенные почки раньше перестали развиваться по сравнению с почками средних и высоких ярусов (табл. 13). Например, почки 5-го узла сорта Димят к 25/VII 1959 г. прорастали всего на 32%, в то время как почки 15-го узла развивались на 94%. Аналогичный вывод сделал также Кондо (1969).
Нuglin (1958) показал также, что наличие точки роста на верхушке побега и летних почек (пасынков) оказывает подавляющее действие на зимующие почки. После удаления этих точек роста они трогаются в рост (рис. 9), причем действие подавления терминальной и летних почек на рост зимующих почек имеет временный характер и не сказывается одновременно на всех глазках по длине побега.
Вопрос о потребности в пониженных температурах в фазе глубокого (органического) покоя, однако, нельзя считать выясненным полностью. Как отмечалось выше, ряд авторов считает, что без воздействия пониженных температур прорастаемость почек не возобновляется. Наиболее четко это понимание выражено в трудах Кондо (1969). Таким образом, воздействие холодом эти авторы рассматривают как средство прекращения периода покоя.
Некоторые авторы придерживаются иной точки зрения. О том, что для прорастания зимующих почек не требуется воздействия пониженной температуры, свидетельствуют данные о развитии винограда в тропических странах. В литературе отмечается, что в условиях тропиков ритм роста и плодоношения винограда осуществляется без воздействия низких температур (Шахова, 1961, и др.). Очень обстоятельный разбор этого ритма сделали Негруль и Мохова (1964), ввиду чего мы не будем распространяться по этому вопросу. Отметим лишь, что в тропических условиях вегетация протекает и заканчивается всегда при высокой температуре, и прорастание почек можно вызвать в любое время.
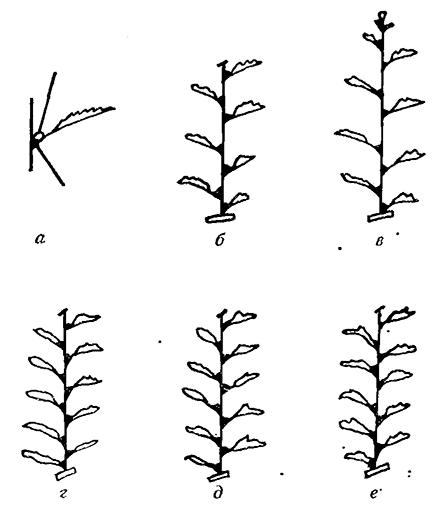
Рис. 9. Корреляция между верхней почкой и летними почками (пасынками), с одной стороны, и зимующими (спящими) — с другой
а — пасынок (спящая почка); б — прищипывание почки; в —пасынкование; г — прищипывание верхушечной почки и пасынкование; д — то же, что и г, но пасынок 10-го яруса сохранился; е— то же, что и д, но пасынки ярусов 6-го и 7-го не удаляются
Негруль и Мохова (1964) показали, что в тепличных условиях без искусственного освещения, но при постоянной высокой температуре можно закончить годовой цикл виноградной лозы примерно за 9 месяцев, осуществляя таким образом 1,2—1,5 вегетаций и периодов покоя в год. 1957), авторы установили состояние покоя и роста почек в любое время года. Конец вегетации наступал всегда, как только заканчивался рост побегов в длину и толщину и начиналось вызревание побегов. Этот переход от одного состояния к другому создавался в процессе эволюции винограда и направлен на подготовку растения к зиме в связи с периодическими переменами погоды. Зимой, как правило, почки находились в состоянии покоя дольше, чем летом. Ритм вегетации осуществлялся также вне зависимости от длины дня.
По современным представлениям, эндогенный покой вызывается накоплением в эмбриональных тканях тормозящих рост веществ (ингибиторов) взамен резкого уменьшения содержания в клетках ауксинов. Природа этих веществ выяснилась после выделения природного ингибитора — абсциссиновой кислоты из плодов хлопчатника. Это дало импульс к изучению покоя почек и семян растений и роли абсциссиновой кислоты в регулировании покоя. Подробные сведения о нем имеются в работе Н. During*.
* См. раздел „Фитогормоны виноградной лозы“ настоящего тома.


Рис. 11. Сопоставление содержания свободной абсциссиновой кислоты (АВА, 10-7 mg на почку) в почках с августа по март 1969/1970 (а) с ходом температуры СО в это же время (б) и с критической продолжительностью (в днях) прорастания сорта Рислинг (в)
1 — сбор урожая ; 2 — наступление холода
Рис. 10. Изменение содержания абсциссиновой кислоты (АВА) в почках винограда во время осенне-зимнего периодаа. Экстинкционная кривая абсциссиновой кислоты (АВА) в экстрактах почек с ноября (1) по декабрь (2) разных проб сорта Рислинг; б. 0 — Д — спектр абсциссиновой кислоты; горизонтальная линия — метанол
Как показали исследования Cheng с сотр. (1974), содержание абсциссиновой кислоты увеличивается с августа, достигая максимума в ноябре. После этого отмечается резкое снижение свободной абсциссиновой кислоты, что совпадает с первыми проявлениями похолодания.
Тем временем увеличивается связанная форма абсциссиновой кислоты (рис. 10, 11). В полной синхронности с этим происходит и прорастание почек.