Из общего объема поглощенной растением воды лишь незначительная доля задерживается в растении, большая часть ее затрачивается на испарение. Транспирация, являясь сложной функцией взаимодействия растения с внешней средой, не только участвует в питании и терморегуляции растений, она не в меньшей мере ответственна за поддержание тургора клеток и за газообмен тканей с атмосферой.
При изучении в полевой обстановке процесса транспирации у многолетников приходится сталкиваться с трудностями методического порядка. Весьма точный весовой метод определения интенсивности транспирации, разработанный применительно к растениям, выращенным в вегетационных сосудах, в данном случае совершенно неприменим. Кобальтовый метод (Stahl, 1894) является скорее качественным. Довольно широкое применение в физиологических определениях получил метод, предложенный независимо друг от друга Ивановым (1918) и Huber (1927) и в дальнейшем усовершенствованный Рихтером и Страховым (1929) и Васильевым (1931).
Многочисленные определения интенсивности транспирации у винограда, выполненные разными исследователями и в различных условиях, показали большую изменчивость данного процесса в зависимости от самых разнообразных причин.
Метамерные, дневные и сезонные изменения интенсивности транспирации
Активность транспирационного процесса неодинакова у листьев, расположенных в разных частях побега. Оникийчук (1962) отмечает, что как в начале цветения, так и в период созревания ягод транспирация у листьев, расположенных выше на побеге, более интенсивная.
Сопоставление по силе испарения листьев различных ярусов показало, что у физиологически более активных и обычно лучше освещенных верхних листьев расход воды в процессе транспирации выше, чем у листьев среднего и тем более нижнего ярусов. По-видимому, этими же причинами следует объяснить и повышенную транспирацию пасынковых листьев, которая по данным Стоева с сотр. (1952) была вдвое выше, чем у листьев основного побега в нижнем и среднем ярусах.
Аналогичная зависимость интенсивности транспирации от принадлежности листьев к пасынковому или основному побегам установлена и в исследованиях Кондо (1960).
Виноградному растению присуща высокая транспирационная активность, хотя проявление ее зависит от многих причин внутреннего и внешнего характера. По данным Geisler(1961) величина транспирации у сорта Рислинг достигает 19—20 mg воды в пересчете на 100 cm2 листовой поверхности за одну минуту. У других сортов он указывает более низкие показатели— 15 mg воды на 100 cm2/min. Contardi и Pimenides (1949, 1950) для условий Аргентины приводят значения транспирации в пределах 10 g на 100 cm2 листовой поверхности за 24 часа.
Согласно Kozma Pal (1954) виноград, культивируемый на сухих песчаных почвах Венгрии, испаряет в середине лета в зависимости от погодных условий 200—1000 g воды в сутки в расчете на один куст.
У сорта Рислинг в Германии куст со 150—200 листьями выделяет при температуре 24 °С от 1 до 1,5 1 воды в сутки.
В условиях жаркого климата Средней Азии потери воды одним кустом винограда на участках, хорошо обеспеченных почвенной влагой, могут превышать 2 1 в час (Кондо, Пудрикова, 1969).
Столь широкий диапазон величин транспирации у винограда обусловлен многими причинами: различиями в климате разных местностей, метеорологическими особенностями отдельных лет, своеобразными погодными условиями в каждом случае определения, запасами продуктивной влаги в почве, физиологическим состоянием растений, фазой их развития, возрастом, сортом и пр.
Определения динамики транспирации у плодоносящих кустов и у молодых растений в школке, выполненные в Молдавии, показали, что в начальный период вегетации интенсивность транспирации в расчете на единицу веса или площади листьев наиболее высока. Однако вследствие еще слабой облиственности растений в это время суммарный для всего куста расход воды оказывается меньшим, чем в другие сроки вегетационного периода.
У взрослых растений наибольшие на протяжении сезона потери воды кустом приходятся на середину лета, в сентябре уже отмечается устойчивое снижение интенсивности транспирации и уменьшение листовой массы. У молодых растений в школке вследствие поздних сроков их посадки (вторая половина мая) вегетация несколько сдвигается, и в сентябре наблюдается нс только довольно активное испарение воды листьями, но и продолжающееся наращивание ассимиляционной поверхности. Поэтому у саженцев в школке расход воды в осенние сроки в расчете на целое растение нередко бывает близким к летнему уровню или даже превышающим его.
Довольно существенно разнится динамика транспирационной активности по годам, что в основном обусловлено метеорологическими особенностями каждого из них. Наблюдаемые в течение дня колебания интенсивности транспирации также в значительной мере вызываются изменениями внешних факторов.
Движение воды в системе растение — атмосфера в известной мере определяется градиентом активности воды между ними (Алексеев, 1948; Гусев, 1966). Естественно поэтому, что изменение условий влажности на любом конце этой цепи неизбежно приведет к усилению или ослаблению транспирационного тока.
Значение для транспирации растений насыщенности воздуха водяными парами отмечалось давно (Максимов, 1926; Алексеев, 1948; Крафте, 1951; Сабинин, 1955). По мнению Негр у ля (1959) недостаточная влажность атмосферы приводит к смыканию устьиц и ограничению транспирации. Коzma Pal (1954), изучавший испарение винограда с помощью специального шкафа, позволявшего измерять всю выделенную кустом воду, устанавливает прямую зависимость интенсивности транспирации от температуры воздуха, дефицита атмосферной влажности и других метеорологических факторов (рис. 5). Как видно, количество выделенной за час воды на протяжении дня сильно меняется. Максимума испарение обычно достигает в околополуденные часы, когда температура и дефицит влажности воздуха наиболее выражены. Изменение внешних условий может привести к смещению максимума.
Среди всех внешних факторов, определяющих активность транспирации, Коzma Pal (1954) выделяет температуру воздуха.
По его материалам ход транспирации, представленный средними за день величинами, находится в почти полном соответствии с изменениями температуры, т. е. чем она выше, тем больше воды испаряет куст, снижение температуры вызывает ограничение данного процесса (рис. 6).

Рис. 5. Суточные изменения транспирации сорта Траминер
1— температура воздуха (°С); 2 — транспирация с 1 куста в g/h;3— дефицит воздушной влажности (%)

Рис. 6. Сезонные изменения транспирации сорта Траминер
1 — среднесуточная температура; 2 — дефицит воздушной влажности;
3 — транспирация с g/h с 1 куста
Такая обусловленность интенсивности транспирации исключительно температурным фактором может иметь место в определенных случаях, например, при небольшом колебании влажности воздуха и при достаточной влагообеспеченности кустов, и, вероятно, только у взрослых растений. Во всех других случаях ее зависимость более многогранна.
Дневной максимум транспирации не всегда совпадает с максимумом температуры, хотя до какого-то момента их нарастание идет почти параллельно (рис. 7). И тем более не всегда в дни с напряженным температурным режимом наблюдается интенсивная транспирация.
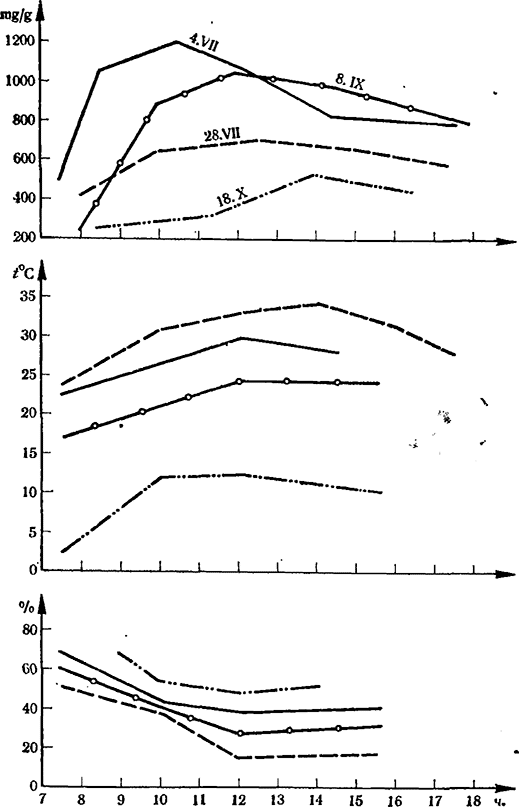
Рис. 7. Динамика интенсивности транспирации листьев (mg/g на час) сорта Ркацители (подвой Рипария х Рупестрис 101-14,1972 г.) в зависимости от температуры воздуха (°С) и относительной влажности воздуха (%)
Сопоставление скорости транспирационного тока в дни, отличающиеся гидротермическими условиями, выявляет большую зависимость этого процесса от совокупности температуры и влажности среды (Пудрикова, 1977). Сравнительно высокая температура (30°С) в середине июля 1972 г., сопровождаемая относительной влажностью воздуха в 40%, способствовала интенсивному испарению листьев, дневной максимум достигал 1200 mg/g на час. Большее напряжение температуры (35°С) и очень низкая влажность воздуха (ОВВ — 18 %) во время атмосферной засухи в конце месяца ограничивали транспирацию. Дневной ход не был представлен однообразной кривой с максимумом в 700 mg/g на час. Сочетание более умеренной и обычной для лета и начала осени температуры в 25°С и существенной сухости воздуха (ОВВ — 30 %) обусловили в начале сентября тот уровень транспирации, который оказался ниже, чем в середине июля, и выше, чем во время атмосферной засухи в конце его.
Следовательно, молодые растения винограда в школке особенно чувствительны к сухости воздуха, ибо их корневая система (к этому времени еще недостаточно развитая) не в состоянии обеспечить листья тем количеством воды, которое необходимо для противоборства атмосферной засухе. Затрудненное поступление воды в листья в этом случае приводит к такому же результату, как и при недостаточном водоснабжении растений при дефиците почвенной влаги, а именно, к усилению водоудерживающей способности тканей и сокращению их транспирации.
Самая низкая транспирация отмечена в конце периода вегетации, когда, с одной стороны, физиологическая активность листьев уже ослаблена и, с другой стороны, пониженные температуры среды не благоприятствуют прохождению этого процесса.
К указанным агентам внешней среды, оказывающим влияние на интенсивность транспирации, следует добавить также силу ветра и солнечную радиацию. Последняя, согласно Максимову (1926), определяет суточный ход почти всех метеорологических факторов и тем самым прямо или косвенно влияет на транспирационную активность. Сабинин (1955) считает свет одним из основных условий транспирационного процесса, рассматривая его как фактор, повышающий проницаемость протоплазмы для воды и регулирующий открытие устьиц.
О большом влиянии света на указанный процесс говорят данные по изменению его активности на протяжении дня: до восхода солнца и после его захода количество испарившейся воды минимально, в часы же наибольшей инсоляции оно выше 2 g в расчете на dm2 листовой поверхности за час (табл. 5).
Таблица 5
Оводненность и интенсивность транспирации освещенных (1) и затененных (2) листьев винограда

Сравнение интенсивности испарения у освещенных и затененных листьев выделило первые из них, отличавшиеся большим испарением и меньшим содержанием в тканях воды.
В природных условиях растения подвергаются воздействию комплекса факторов, первое место среди которых занимает содержание продуктивной влаги в почве, так как достаточное количество последней может ослабить, а дефицит — усилить угнетающее действие каждого из них. Поэтому в обстановке, в которой растения не испытывают острого недостатка в водоснабжении, интенсивность транспирации в основном следует за изменениями температуры и влажности воздуха.
Исследования, выполненные И. Н. Кондо в Средней Азии, показали, что на участках с хорошей обеспеченностью растений почвенной влагой интенсивность транспирации весьма велика в течение всего дня, максимум ее обычно приходится на часы наибольшего напряжения атмосферных факторов и достигает двух и более граммов воды на 1 g свежего веса листьев в час.
На неорошаемом винограднике (в зоне необеспеченной богары) активность транспирационного процесса в тот период, когда содержание влаги в корнеобитаемых слоях почвы значительно выше коэффициента завядания, также довольно высока. Во второй половине лета виноградные кусты на богаре, испытывающие воздействие сильной засухи, расходуют воду очень экономно; интенсивность транспирации резко падает (почти в 4—10 раз); максимальная трата воды приходится на утренние часы (Кондо, Пудрикова, 1969).
Таблица 6
Влияние подвоя и метеорологических условий различных лет (1965, 1966, 1968, 1970) на интенсивность транспирации винограда

* Обозначения степени достоверности те же, что и в табл. 2.
Как видим, рассмотрение всех отдельных случаев проявления транспирационной активности винограда показывает большую зависимость ее от факторов внешней среды. Указанная связь подтверждается результатами дисперсионного анализа.
Сравнение по данному признаку растений двух сортов — Алиготе и Шасла, — привитых на различных подвоях, в аналогичные сроки разных лет показало, что влияние метеорологических особенностей этих лет является наиболее весомым — сила воздействия их оценивалась в общей сумме зависимостей 59—89% у Алиготе и 66—82% у Шаслы (табл. 6). Влияние сортовой специфики подвоев было менее существенно: 4—9% у первого сорта и 1-6 % — у второго.