II.2. МОРФОФИЗИОЛОГИЧЕСКИЕ ФАКТОРЫ ФОРМИРОВАНИЯ КРОНЫ ВИНОГРАДА КАК ПРОДУКЦИОННОЙ СИСТЕМЫ
Изменение архитектуры и продуктивности кроны растений как оптико-биологической системы зависит от ритмичности физиологических процессов, связанных с развитием побегов и работой листового аппарата. Определяющим в этом процессе является видовая специфика растения и генетическая приспособленность его к условиям среды.
Рост побегов винограда начинается после распускания почек и в зависимости от сорта и условий культуры продолжается до начала вызревания побегов или даже до наступления полной зрелости ягод. Наиболее интенсивно побеги обычно растут в период от распускания почек до начала цветения. К концу этого периода рост их достигает 60% и больше полной длины, а суточный прирост —8—10 см [200, 236]. Снижение скорости роста побегов в период цветения не является обязательным. При некоторых условиях скорость роста возрастает во время цветения или же после его окончания и достигает 5— 10 см в день [245, 483].
Междоузлие над листом растет в длину в течение 18— 20 дней и достигает максимального прироста, когда становится четвертым—седьмым; рост приостанавливается у восьмого—десятого междоузлия, считая от верхушки побега. В период цветения из сформировавшихся 19 листьев заканчивают свой рост 7—8 от основания побега, а 12 листьев достигают необходимой величины, чтобы отдавать ассимиляты другим частям растения [355].
У значительного числа сортов к моменту цветения побеги имеют по 14—16 узлов с листьями [170]. Удлинение побега происходит в основном путем вытягивания образовавшихся молодых клеток междоузлий (интеркалярный рост) и только отчасти за счет деления клеток верхушечной меристемы. На каждом побеге зона наиболее интенсивного вытягивания охватывает второе, третье и четвертое междоузлия, считая от верхушки. Рост нижележащих междоузлий постепенно замедляется и примерно на шестом-седьмом междоузлии совершенно прекращается. Одновременно с удлинением побега происходит его утолщение: в верхней зоне только за счет увеличения размеров клеток, а ниже — в результате возникновения вторичной меристемы (камбия), формирующей новые концентрические слои флоэмы и ксилемы. Одновременно с ростом побегов увеличивается количество листьев на побеге, их размер, изменяется облиственность побега и всего куста.
Рост листа начинается с вытягивания бугорка на верхушке побега в удлиненную зачаточную пластинку листа. Пластинка растет своим основанием по направлению к верхнему концу.
Общая продолжительность роста листа в среднем 4—5 недель, В течение 6—8 дней пластинка растет медленно, и среднесуточный прирост ее составляет 2—8 см2. Наибольший прирост пластинки происходит во вторую и третью недели — до 14—20 см2 в день. К концу периода среднесуточный рост пластинки снижается до 3—5 см2. Рост пластинки останавливается, когда лист становится 12—16-тым от верхушки побега. Независима от сорта, увеличение площади листьев винограда имеет вид сигмоидальной кривой, т. е. большой кривой роста [461, 483]. Площадь листовой пластинки в момент завершения роста в зависимости от сорта составляет 120— 300 см2 и более. Наибольшими размерами характеризуются листья среднего яруса (9—12-тый), наименьшими — верхние листья (21 — 25-тый). Нижние листья (1—5-тый) имеют пластинку промежуточной величины.
В течение первых 30— 40 дней после появления листьев фотосинтетическая активность единицы площади листа повышается и к концу указанного срока достигает максимума [454]. В последующие периоды вегетации наблюдается общая тенденция постепенного снижения фотосинтеза листьев [220, 285, 339, 481]. По Кридеману [454] (рис. 3), кривая фотосинтеза винограда в зависимости от возраста листа описывается уравнением, имеющим вид обратного полинома:
х/у= В0 + В1х+В2х2,
где х — средний возраст листа (дни), у— фотосинтез (мг СO2/(дм2-ч)), В0 = 2,249, В1 = 0,048, В2 = 0,0011227.
В период вегетации у винограда отчетливо проявляется изменение фотосинтетической активности листьев, расположенных на разных ярусах побега. В начале вегетации выше активность листьев нижнего яруса (5—12-тый узлы побега). В середине и в конце вегетации зона высокой фотосинтетической активности перемещается к листьям среднего яруса (15—20-тый узлы) и верхней части побега [76, 231, 241, 332, 338].

Рис. 3. Фотосинтез (1) и фиксация 14СО3 (2) в зависимости от возраста листа (числа дней от распускания почек) (по [454]).
Отток ассимилятов из листьев начинается, когда листья достигают около половины своего конечного размера. У молодых 2—3-недельных побегов длиной около 15 см листья еще не поставляют продукты фотосинтеза, и в этот период побеги потребляют пластические вещества, поступающие из других частей куста [435, 461].

Рис. 4. Главные направления передвижения ассимилятов в побегах винограда в зависимости от фазы развития растения.
А — в очень молодых побегах, Б — в период цветения, В — после завязывания ягод (по [435]).
При дальнейшем развитии побега листья начинают поставлять ассимилянты главным образом в акропетальном направлении, к верхушке побега (рис. 4 А) [435]. Такое направление передвижения ассимилятов продолжается не более одного-двух дней. С появлением следующего листа часть продуктов начинает передвигаться базипетально в направлении к гроздям и. многолетним частям куста. Двустороннее передвижение ассимилятов продолжается 2—3 дня, после чего направление передвижения ассимилятов полностью становится базипетальным . Прежнюю функцию листьев — снабжение растущей верхушки-побега продуктами фотосинтеза — принимают на себя более молодые листья, расположенные в верхней части побега. Такое распределение ассимилятов характерно для периода до цветения и в период цветения, когда направление передвижения-ассимилятов из листьев молодого побега определяется двумя-потребителями: верхушкой побега и многолетними частями. Соцветия в этот период являются относительно слабыми потребителями ассимилятов. Незначительное количество ассимилятов поступает в соцветия из листьев, расположенных выше соцветия (рис. 4 Б). Слабая конкурентная способность соцветий винограда как потребителей до момента завязывания ягод показана также Кумбом [423].
С момента окончания цветения (завязывания ягод) и на протяжении всего периода созревания передвижение ассимилятов из листьев, расположенных ниже грозди, осуществляется в двух направлениях: к грозди и к основанию куста. Акропетальное передвижение из этих листьев не распространяется выше грозди. Листья, расположенные над гроздью, также поставляют ассимиляты в двух направлениях: к грозди и к верхушке побега (рис. 4 В). Роль верхушки как потребителя в связи с уменьшением энергии роста в этот период ослабевает. В начале созревания и в период полной зрелости винограда ассимиляты из листьев всех ярусов, включая и расположенные в 10 см от верхушки побега, направляются к гроздям и многолетним частям растения.
Помимо передвижения к основным потребителям (верхушка побега, грозди, многолетние части), ассимиляты в побеге передвигаются и к так называемым «второстепенным» потребителям (глазки, пасынковые побеги, усики, камбий и др.). В отличие от главных, «второстепенные» потребители не оказывают влияния на продольное направление передвижения ассимилятов в побеге.
Передвижение ассимилятов из листа имеет характер «линейности», т. е. оно ограничено вертикальной транспортной системой более или менее в линии с листом. При определенных условиях линейность передвижения изменяется в результате-действии главных потребителей. Помимо линейного передвижения, имеет место поперечное, или латеральное, распределение ассимилятов, главным образом в верхушечной части побега.
Чанишвили [390] отмечает наиболее интенсивный отток ассимилятов из листьев среднего яруса. В восходящем направлении ассимиляты передвигаются со скоростью 33—44 см/ч, в нисходящем—15—17 см/ч. В опытах Дурмишидзе [117] скорость восходящего тока ацетата, меченного по углероду, в срезанных стеблях винограда на свету составила 80 см/ч, а нисходящего— 6 см/ч. Согласно Соунсону [333], скорость транспорта сахарозы в виноградном растении составляет 1 см/мин. Скорость передвижения в нисходящем направлении значительно меньше.
Как известно, у некоторых растений существует автономность транспортных связей между листьями и другими частями растения. Так, например, у подсолнечника в период формирования семян каждый лист снабжает ассимилятами определенный сектор корзинки. У корнеплодных растений (редька, морковь, свекла) каждый взрослый лист снабжает продуктами фотосинтеза только определенную часть корнеплода и прилегающие к ней молодые листья [79, 391]. Исследования показали [281], что у виноградного растения строгая автономность транспортных связей между листьями и другими органами растения отсутствует. Распределение продуктов фотосинтеза из листьев разных ярусов отличается широким захватом всех органов побега. Ассимиляты поступают из меченого листа в соседние на стрелке побеги, в побеги на противоположной части куста и в корни. Вместе с тем отток и распределение продуктов фотосинтеза из листьев разных ярусов изменяется по периодам вегетации: по мере роста побегов и нарастания новых листьев зона наиболее продуктивных листьев перемещается от нижних к верхним ярусам. Негруль и Никифорова [238] выявили функциональную взаимосвязь между отдельными частями побега с помощью метода кольцевания. В опытах Стоева [335] в секторе третьего и седьмого узлов ассимиляты из подкормленного 14СО2 листа не поступали в соседние листья. В [435] наблюдали это только в том случае, когда подкормку проводили в листьях, расположенных на 6—7 узлах от верхушки побега.
После окончания цветения суточные приросты побегов снижаются, хотя затухающий рост побегов может продолжаться значительное время. По наблюдениям Потапенко [291], в районе Новочеркасска во второй половине июля из 225 сортов винограда первые признаки окончания роста побегов отмечались только у 20 сортов; на 17 августа рост побегов продолжался почти у 50 сортов.
Особенностью винограда является неравномерность развития побегов. На рис. 5 в виде гистограмм показан характер развития побегов у двух сортов, различающихся по силе роста: сильнорослого сорта Хиндогны и среднерослого сорта Мускат белый.
У Муската белого кривая распределения длины побегов во все сроки заметно отклоняется от нормального и приближается к асимметричному. Значительный удельный вес в выборках занимают слаборослые побеги (крайние левые классы). У сорта Хиндогны кривые распределения в начале вегетации приближаются к нормальному (29 мая, 20 июня), но затем по мере роста побегов начинает проявляться асимметрия ряда с эксцессами, вершины которых выступают за пределы нормального распределения. В целом у обоих сортов (в большей степени у Муската белого) асимметрию в распределение вносит группа отстающих в развитии побегов крайней левой части вариационных рядов.
В табл. 1 показаны числовые статистические характеристики.

Рис. 5. Гистограммы распределения длины побегов в разные периоды вегетации у сортов Мускат белый (слева) и Хиндогны (справа). Южный берег Крыма, 1969 г. (объем выборки «=180).
Естественный разброс изучаемого материала характеризуют значения среднего квадратического отклонения δ, коэффициента вариации V и коэффициента асимметрии А.
Таблица 1
Статистические характеристики длины побегов у сортов винограда на Южном берегу Крыма (1969 г.)
Дата | Размах варьирования, см | M*см | δ см | v % | A |
| Мускат белый | ||||
29 V | 11—77 | 31 | 21 | 38,7 | -3,15 |
20 VI | 27—202 | 89 | 43 | 38,3 | -1,09 |
15 VII | 27—273 | 112 | 69 | 61,6 | -1,47 |
12 VIII | 27—294 | 114 | 72 | 63,1 | -1,14 |
| Хиндогны | ||||
29 V | 18-73 | 34 | 12 | 35,2 | —0,72 |
20 VI | 31—193 | 100 | 34 | 34,0 | —1,74 |
15 VII | 35—281 | 134 | 55 | 41,0 | — 1,49 |
22 VIII | 36—298 | 140 | 59 | 42,1 | —1,49 |
Высокие показатели коэффициента вариации (до 65,3% у Муската белого и 44,6% у Хиндогны) свидетельствуют о большой пестроте и неоднородности материала. У обоих сортов варьирование длины побегов возрастает по мере их роста.
Коэффициенты асимметрии указывают на значительную асимметрию в распределении со смещением средней арифметической влево по оси абсцисс (отрицательный знак коэффициента), что совпадает с графическим представлением вариационных рядов.
На рис. 6 показаны гистограммы распределения длины побегов сорта Ркацители при нормальной и повышенной нагрузках побегами. По мере роста побегов кривая распределения также отклоняется от нормального. Наибольший удельный вес в выборках занимают побеги сравнительно небольшой длины (60—70 см), относящиеся к левым классам. На кустах с повышенной нагрузкой увеличивается доля слаборослых побегов, повышается неоднородность в их развитии, что проявляется в более высоких значениях коэффициента вариации (при последних замерах V = 47% при нормальной нагрузке и 57% при повышенной).
Структура кроны оказывает влияние на соотношение освещенных и затененных листьев. Показано, что при ориентации рядов с севера на юг на вертикальной шпалере кусты имеют до 40% затененных листьев [323]. В зависимости от густоты посадки количество полностью затененных листьев в кроне может увеличиваться от 33 до 50% [243]. В наших опытах у сорта Ркацители при площади листьев кустов 3,7 и 5,3 м2 доля затененных листьев внутри кроны составила соответственно 22 и 25%.
Период окончания роста побегов совпадает с вызреванием побегов — важнейшим физиологическим процессом, определяющим продолжительность жизни виноградного растения и его продуктивность. Вызревание однолетних побегов, как правило, начинается с нижних междоузлий от основания побега и постепенно распространяется вверх. Отмечаются случаи, когда раньше начинают вызревать междоузлия в средней части побега [291, 378].
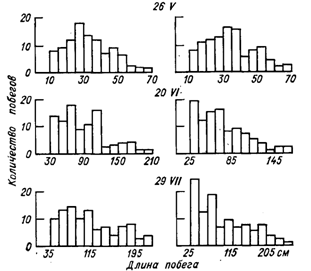
Рис. 6. Гистограммы распределения длины побегов в период вегетации при нагрузке 18 (слева) и 40 побегов на куст (справа). Сорт Ркацители, Западная степная прибрежная зона Крыма, 1972 г. (объем выборки и=160).
Темпы, сроки и степень вызревания зависят от многих факторов. К числу важнейших относится свет. При недостатке его побеги вызревают слабо [150]. Наиболее важными анатомическими изменениями в процессе вызревания побегов являются образование пробковой ткани (перидермы), а также утолщение и одревеснение клеточных стенок в результате накопления в них лигнина, гемицеллюлозы и минеральных веществ. К наиболее важным физиологическим изменениям побега в этот период следует отнести понижение уровня содержания воды и максимальное накопление в запасающих тканях питательных веществ, прежде всего крахмала [150, 284, 335].