Андреасен и Штир (1953) показали, что при введении эргостерина в бродящую среду можно получать, действуя в строго анаэробных условиях, среднюю численность популяции от 110 до 125 млн. клеток на 1 см3, которые накапливаются после большого числа последовательных пересевов в среде, совершенно лишенной кислорода. Оптимальная доза эргостерина равна 7 мг/л. С другой стороны, в анаэробиозе наблюдается небольшой ингибирующий эффект. Авторы пришли к заключению, что эргостерин следует рассматривать как один из необходимых факторов развития дрожжей в условиях анаэробиоза, потому что именно в таких условиях они не способны реализовывать синтез стеринов. Эта потребность также удовлетворяется холестерином, но не всеми стеринами. Эргостерин оказывается также необходимым для развития дрожжей при 40° С, так как его синтез сильно тормозится высокой температурой (Старр и Парке, 1962).
Большой интерес, который представляют эти наблюдения для практики брожения, не ускользнул от внимания исследователей, дающих на них многочисленные ссылки (Можене и Дюпон, 1964; Шове и сотрудники, 1966).
В табл. 8.4 приведены результаты серии экспериментов, проведенных на различных винных дрожжах (Институт энологии, Бордо, 1965), которые полностью подтверждают стимулирующие свойства эргостерина при добавлении его в суспензию (5 мг/л) с Твином-80. Этот продукт действует на все дрожжи. Он обеспечивает более полное сбраживание сахара, и разница в конце брожения составляет от 40 до 150 г сахара. Такое брожение имеет характеристики аэробного процесса: более интенсивное образование глицерина, меньшее образование ацетальдегида и уксусной кислоты.
Роль эргостерина проявляется также при брожении в условиях повышенной температуры: он позволяет, например, дрожжам лучше выдерживать температуру 37° С и при этом образовывать дополнительно около 4% об. спирта. Действие эргостерина тем эффективнее, чем раньше его вводят во время брожения; но даже и при добавлении в конце он может помочь сбродить больше сахара.
Было взято виноградное сусло сахаристостью 260 г/л. Брожение проводили без доступа воздуха первые 3 дня при 25° С и в последующие 15 дней при 37° С.
Количество сброженного сахара (в г/л) приведено ниже.
Контроль 84
4-5 мг/л эргостерина при забраживании 150
+ 5 мг/л эргостерина через 3 дня 136
при 25° С
Таким образом, подтверждается, что эргостерин активирует брожение и рост дрожжей в анаэробиозе и придает этим процессам характеристики, присущие им в условиях аэробиоза. Эргостерин замещает в некотором роде кислород для дрожжей. Состояние анаэробиоза затрудняет синтез эргостерина, так как достаточно добавить это вещество, чтобы смягчить недостаток кислорода.
Поскольку эргостерин нерастворим, его переводили во время этих экспериментов в состояние стабильной суспензии с помощью Твина-80 (частичный сложный эфир олеиновой кислоты и ангидрида сорбита). Этот продукт вызывает помутнение сусел и вин. Будучи растворимым в спирте при высоких температурах, эргостерин осаждается, когда его добавляют в водную среду. Его можно диспергировать в диатомите, но получаемый порошок имеет меньшую активность, чем коллоидная суспензия.
Активностью, аналогичной активности эргостерина, обладают также некоторые стероиды и жирные кислоты с длинной цепью. Некоторые из них входят в состав пруина виноградных ягод, что делает их в свете недавних исследований очень важными для виноделия.
Опыты, проведенные в прошлом, показали неожиданно большое влияние кожицы винограда на анаэробное брожение сусла (табл. 8.5). При этом констатировали, что кожица (мезга) в виде плавающей или погруженной шапки не ускоряла аэробного брожения, но активировала брожение в анаэробных условиях до такой степени, что в присутствии кожицы было сброжено большее количество сахара без доступа воздуха, чем в аэробных условиях. Промывание кожицы водой или разведенным спиртом не уменьшало количества активного вещества.
Таблица 8.5
Влияние кожицы виноградных ягод (Мезги) на брожение сусла
Показатели | Контроль | Количество сахара, сброженного на 5-й тень (в г/л), при доб (влении кожицы винограда (100 г/л) | |
белых сортов | красных сортов | ||
Брожение |
|
|
|
с доступом | 131 | 126 | 124 |
воздуха |
|
|
|
без доступа | 84 | 156 | 140 |
воздуха |
|
|
|
Точно так же и семена винограда несколько активируют брожение и заметно ускоряют рост дрожжей. В одном из опытов популяция дрожжей возросла на 40%, но семена уступают по своему воздействию на ход брожения активаторам сусла.
Влияние экстрактов пруина на размножение дрожжей в анаэробиозе (по данным Брешо и сотрудников, 1966)
Экстракт пруина | Количество дрожжевых клеток через 5 дней (в тыс. на 1 мм3) | ||||
контроль без пруина | при добавлении экстракта пруина (в г/л) | при добавлении эргостерина (25 мг/л) + +Твин-80 | |||
0,37 | 0,75 | 1,50 | |||
Экстракт | 50 | 72 | 83 | 102 | 107 |
Работы Брешо и сотрудников (1966, 1971) должны были дать объяснение этим явлениям. Авторы готовили экстракты пруина из винограда сорта Гаме вымачиванием гроздей в хлороформе или в эфире. После испарения растворителей полученный экстракт пруина растворяли в абсолютном спирте и добавляли в различных дозах в виноградный сок. В табл. 8.6 приведены некоторые результаты этих опытов. Они показывают положительное действие экстрактов пруина, эргостерина и сложного эфира олеиновой кислоты. Брожение сусла сахаристостью 250 мг/л проводилось под азотом.
Следовательно, по мере того как активные вещества пруина становятся при переработке винограда растворимыми, они ведут себя как факторы роста дрожжей. Брешо и сотрудники (1966) видят в этом явлении объяснение факта хорошего роста дрожжей при виноделии с настаиванием без доступа кислорода.
Между тем известно, что некоторые ненасыщенные жирные кислоты, как олеиновая, линолевая (Нордгейм и Рихе, 1966), линоленовая играют в отношении бактерий молочнокислого брожения роль такого же фактора роста, как биотин. Суомалайнен (1965) показал, что в присутствии аспарагиновои кислоты пальмитиновая и олеиновая кислоты также могли бы заменять биотин для Saccharomyces (рис. 8.3).
Наконец, Радлер (1968) в своем исследовании компонентов пруина или кутикулярного воска виноградной ягоды показал, что он наполовину состоит из окситериеновой и олеанолевой кислот.
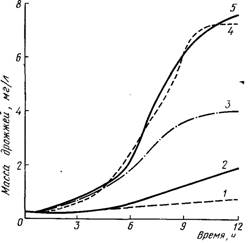
Рис. 8.3. Рост пекарских дрожжей с применением биотина и без него, но с добавлением аспарагиновой и олеиновой кислот:
1 — без биотина; 2— олеиновая кислота; 3—. аспарагиновая кислота; 4 — олеиновая кислота; 5 — биотин.
Другая же половина представлена парафинами, альдегидами, сложными эфирами, спиртами, ненасыщенными и насыщенными жирными кислотами большой молекулярной массы (см. гл. 3, раздел «Состав кожицы»).
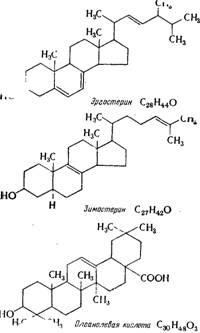
Рис. 8.4. Структура некоторых стероидов, участвующих в росте дрожжей.
Родство структур олеанолевой кислоты и стеринов дрожжей приведено на рис. 8.4.
Содержание этой кислоты и ненасыщенных жирных кислот в пруине заставили Брешо и сотрудников (1971) поставить перед собой вопрос, не играет ли этот стероид, один или вместе с олеиновой кислотой, какую-то роль в анаэробном росте дрожжей.
Табл. 8.7 составлена по результатам экспериментов этих авторов. Она позволяет измерять соответствующую эффективность этих различных факторов. Олеанолевая кислота оказывает неоспоримое стимулирующее действие, даже большее, чем у эргостерина. То же самое можно сказать и о смеси олеанолевой и олеиновой кислот при сравнении со смесью эргостерина и олеиновой кислоты. В присутствии стероидов число 'клеток удваивается или утраивается по отношению к анаэробному контролю и может даже превосходить значения аэробного контроля.
Эти результаты показывают, что олеанолевая кислота, одна или совместно с олеиновой кислотой, способствует росту дрожжей в анаэробных условиях, несмотря на почти полную нерастворимость этого вещества. Этим объясняются стимулирующие свойства пруина винограда в анаэробных условиях, обычно имеющих место при некоторых способах виноделия. Они также могли бы, наверное, объяснить скорость брожения и свою удивительную эволюцию при повышенной температуре в процессах переработки винограда с применением термической обработки перед дроблением.