Рассмотрение запаса витаминов в вине по сравнению с суслом ясно показывает, что в результате брожения не уменьшается содержание витаминов в среде, но происходит глубокая перестройка витаминного состава. Женевуа и Флавье (1939) впервые указали на эту эволюцию; они наблюдали, что в ходе брожения тиамин быстро убывает, тогда как рибофлавин накапливается. Другие авторы проследили эволюцию витаминов группы В от винограда до вина (Кастор, 1953; Холл и сотрудники, 1956). Пейно и Лафон-Лафуркад (1957) детально описали эти превращения. Рис. 8.1 воспроизводит поведение некоторых факторов роста.
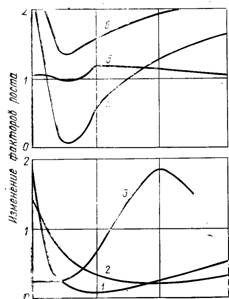
Рис. 8.1. Эволюция основных факторов роста во время сбраживания виноградного сусла дрожжами Sacch. ellipsoideus:
1 — тиамин (100 мкг); 2— биотин (1 мкг); 3 —рибофлавин (100 мкг), 4 — никотинамид (1 мг); 5 — пантотеновая кислота (1 мг); 6 — пиридоксин (1 мг).
Пантотеновая кислота изменяется во время брожения по-разному в зависимости от вида дрожжей, на которых проводят эксперимент. В первые часы Sacch. ellipsoideus синтезирует ее или высвобождает из своих связанных форм. Затем процентное содержание этого витамина быстро снижается в период логарифмического размножения клеток; наконец, оно возрастает, по всей видимости, в результате выделения. Другие прототрофные дрожжи ведут себя таким же образом. Наблюдают непрерывное поглощение пантотеновой кислоты ауксотрофными дрожжами, такими, как Kl. apiculata. В виноделии обычно констатируют увеличение содержания этой кислоты, и красные вина несколько богаче ею, чем вина белые. Во время выдержки наблюдается медленное разрушение пантотеновой кислоты.
Kl. apiculata и Sacch. rosei вызывают почти полное исчезновение пиридоксина во время брожения, и, наоборот, другие дрожжи, такие, как С. pulcherrima и Т. stellata, удваивают или учетверяют процентное содержание пиридоксина в среде. Sacch. ellipsoideus поглощают значительное количество пиридоксина в стадии быстрого размножения клеток; затем его содержание в бродящей среде прогрессивно возрастает, заметно приближаясь к исходному уровню. В нормальных условиях приготовления вина эти модификации завершаются выравниванием содержания пиридоксина в сусле и винах. Белые или красные вина содержат его в очень близких дозах. Длительное хранение вина не оказывает влияния на пиридоксин.
Все дрожжи поглощают значительные количества биотина, особенно в условиях аэробиоза, во время фазы размножения. На 5-й день 9/10 первоначального количества биотина в сусле исчезали. Концентрация биотина возрастает вследствие выделения, и в конечном счете, в условиях производства вина практически содержат такое же количество биотина, как и сусло, из которого они изготовляются.
Никотинамид широко используется в сусле всеми дрожжами. Sacch. ellipsoideus снижают его содержание в среде уже на 3-й день с 2,2 до 0,1 мг. В дальнейшем процентное содержание никотинамида в жидкости возрастает, но не достигает первоначального уровня. Этот фактор роста необходим тем более, чем более анаэробными являются условия культуры (Суомалайнен и сотрудники, 1965). Вино обычно беднее никотинамидом, чем сусло, из которого оно изготовлено. Красные вина имеют более высокое содержание никотинамида, чем белые. Впоследствии никотинамид становится стабильным витамином.
Потребности дрожжей в тиамине очень различны в зависимости от их вида, в то время как некоторые, например Sacch. ellipsoideus, Kl. apiculata, Sacch. rosei, практически истощают среду (первоначальная концентрация 330 мкг снижается до 18), другие дрожжи, как P. ferraentans, синтезируют значительные дозы тиамина. Однако первые имеют большее распространение в производстве. В конце концов, брожение завершается исчерпанием тиамина в бродящей среде; в белых винах остаются лишь незначительные дозы его. Максимальное количество тиамина, которое могут поглощать Sacch. ellipsoideus, составляет от 600 до 800 мкг/л. В суслах не всегда бывают такие большие концентрации; понятно, что нередко добавление тиамина могло бы оказывать благоприятное влияние на ход спиртового брожения.
Урнак (1970) исследовал кинетику выделения тиамина; необнаруживаемый в первые дни, он выделяется в течение 3 мес, следовательно, выпадение дрожжевого осадка и снятие с дрожжей прерывают этот процесс, как правило, до того, как будет выделен весь тиамин дрожжей.
Мезоинозит не претерпевает заметных изменений в период брожения, поскольку потребности дрожжей в нем меньше чем 0,5 мг/л, а в сусле содержится в тысячи раз больше мезоинозита, чем необходимо. Белые вина богаче им, чем красные; последние могут подтвергаться молочнокислому брожению, бактерии которого потребляют мезоинозит; некоторые красные вина лишены его.
Цианкобаламин, или витамин B12, может быть обнаружен в винах в дозах порядка нескольких десятых микрограмм на 1 л. Присутствие в винах следов этого витамина можно объяснить способностью дрожжей синтезировать очень малые количества веществ, обладающих активностью В12.
В первой фазе брожения наблюдают значительный синтез парааминобензойной кислоты дрожжами. В частности, Sacch. rosei доводят за 48 ч ее содержание в среде с 25 до 175 мкг/л, и, наоборот, к концу брожения дрожжи, и это относится к Sacch. ellipsoideus, поглощают эту кислоту. Kl. apiculata проявляет себя непрерывным образователем этого витамина.
Что касается холина, то после увеличения на несколько миллиграммов в начале брожения отмечается расходование при снижении его в вине до 2/3 первоначального количества, имевшегося в сусле. С заостренными дрожжами расходование происходит слабо. Как и другие витамины, холин выделяется дрожжами. В практике виноделия брожение сусла характеризуется незначительным потреблением холина и интенсивным образованием парааминобензойной кислоты.
Содержание рибофлавина в сусле невелико, от 3 до 60 мкг/л. Брожение всегда сопровождается синтезом этого витамина, более значительным в аэрированной среде. Синтез протекает энергичнее с окислительными дрожжами, образующими пленку на поверхности питательной среды. В сусле, которое сбраживается обычной микрофлорой, уже со второго дня наблюдают возрастание процентного содержания рибофлавина. Оно достигает максимума в период между пятым и десятым днем. В дальнейшем процентное содержание рибофлавина понижается. Условия приготовления вина мало изменяют его уровень. Красные и белые вина также имеют в своем составе рибофлавин (до 240 мкг/л). Но его большая светочувствительность является одной из причин последующего снижения содержания в белых винах этого фактора роста бактерий молочнокислого брожения.
В конечном счете спиртовое брожение перестраивает и даже дополняет витаминный состав сусла. В действительности это хороший способ сохранения витаминных элементов плода. Прежде всего, отмечается контраст между изменениями тиамина и рибофлавина: когда в сусле много тиамина и мало рибофлавина, вино богато рибофлавином и практически лишено тиамина. Содержание никотинамида в белых винах составляет только 60% от содержания его в сусле, но в красных винах он находится в таких же количествах, как и в винограде. Пантотеновая кислота, пиридоксин, бнотин используются и затем выделяются дрожжами, вследствие чего в конечном счете они находятся в таком же количестве в вине после брожения, как и перед его началом; мезоинозит практически не изменяется.
Запас витаминов в молодом вине может со временем уменьшаться. Воздействие работы бактерий, обработки для осветления, экспортирование на свету, длительная выдержка могут разрушить некоторые витамины полностью или частично. В то время как пантотеновая кислота, иикотинамид, пиридоксин проявляют достаточную стабильность, биотин и особенно рибофлавин могут почти полностью исчезать.
Потребности дрожжей в факторах роста
Если некоторые факторы роста иногда и содержатся в вине в недостаточном количестве, оно никогда не бывает полностью лишено ни одного из основных факторов. И соответственно, если допустить, что в некоторых случаях вследствие частичной недостаточности того или иного фактора вино становится менее восприимчивым к воздействию микроорганизмов, то совершенно невозможно реализовать хорошую биологическую стабильность истощением того или иного необходимого фактора. В действительности нужно учитывать тот факт, что многие микроорганизмы вина и в особенности дрожжи являются прототрофами, т. е. они способны в определенной мере синтезировать недостающий фактор.
Здесь умышленно не рассматриваются качественные потребности дрожжей. Этот вопрос был описан в гл. 5, а примеры приводились в гл. 6. Эти потребности были также исследованы в многочисленных работах (Верона и Пиччи, 1954; Викен и сотрудники, 1954; Викен и Агте, 1958; Вайнфуртнер и сотрудники, 1959; Виндиш и сотрудники, 1967). Было также показано, что потребности дрожжей в факторах роста могут изменяться в зависимости от температуры и степени аэрации (Бич, 1969).