Установлено, что виноградное растение подобно многим древесным породам при благоприятных условиях закаливания в потенции способно развивать более высокую морозоустойчивость, чем это проявляется в природе (Погосян, 1967 а, 1975).Следовательно, гибель от мороза нельзя объяснить только факторами, предшествующими зимнему периоду, или снижением температуры критического уровня, характерного для данного вида или сорта. Устойчивость к отрицательным температурам проявляется в способности растений к соответствующей структурной и функциональной перестройке в изменившихся условиях.
Одной из важных особенностей такой перестройки является приспособительная реакция растений к скорости замораживания, образования в растительных тканях льда, к быстроте процесса оттаивания, т. е. к физиологическим процессам, имеющим часто решающее значение для устойчивости против критических температур.
Исследования с применением быстрого охлаждения черенков винограда, в частности, в области критических температур, а также с регистрацией динамики замерзания тканей (Р ogosian, Sakai, 1969; Погосян, 1971, 1972) показали, что закаленные растения морозостойких видов винограда (V. amurensis, V. coignetiae Pull.), обладающих высокой устойчивостью к замораживанию при сравнительно медленном снижении температуры (со скоростью 3—4° в минуту, близки к естественным суровым условиям), относительно устойчивы и к быстрому охлаждению со скоростью 30—50° в минуту. Растения слабоустойчивых сортов V. vinifera в аналогичных условиях быстрого охлаждения проявляют высокую чувствительность, полностью погибая даже при —14°.
Устойчивость к быстрому охлаждению значительно повышается после кратковременного замораживания при —5 и —10° (по одному часу); при этом растения морозостойких видов выдерживают —23—28°, а слабоустойчивые с большими повреждениями —17—18°. Различия в устойчивости тканей лозы к одним и тем же отрицательным температурам, но при различной скорости охлаждения объясняются быстротой понижения температуры в них
в процессе замораживания. При медленном охлаждении (3—4° в минуту) замерзание клеточного сока обычно начинается при —4—6° (время переохлаждения составляет 60—100 s), а при быстром промораживании — при —11—12° с переохлаждением в течение 3—8 s, в то время как точка замерзания клеточного сока в зимующих тканях винограда составляет —1,9—2,6°. Следовательно, продолжительность переохлаждения тканей при медленном заморживании примерно в 15—20 раз превышает продолжительность переохлаждения при быстром замораживании. При скорости охлаждения 30— 50° в минуту льдообразование начинается в межклетниках, но кристаллы льда легко и быстро (за 10—15 s) могут проникать и в клетку, вызывая внутриклеточное замерзание и ее гибель (Sakai, Оtzukа, 1967; Красавцев, 1972).
Предварительное замораживание черенков при —5 и —10° способствует уже некоторому обезвоживанию клетки, причем период переохлаждения составляет 25—40 s, а минимальная точка переохлаждения доходит до —7, —9°, обеспечивая тем самым лучшую приспособляемость тканей к последующему быстрому охлаждению.
При медленном охлаждении лед образуется внеклеточный, и когда охлаждение достигает критических температур, клетки уже бывают обезвоженными в определенной степени, достаточной для выживания при краткосрочном действии критической для данного индивидуума температуры.
Следовательно, устойчивость виноградной лозы к быстрому охлаждению достигается благодаря своевременному оттоку внутриклеточной воды в межклетники и способности протопласта препятствовать льдообразованию внутри клеток.
Выживаемость тканей виноградной лозы в значительной степени зависит от скорости оттаивания, что в свою очередь обусловлено температурой и теплопроводностью среды, в которой протекает этот процесс (Lоvе1ак, 1954; Туманов, Красавцев, 1962; Sakai, Yoshida, 1967).
При быстром оттаивании (со скоростью от 35 до 180°/min) важное значение имеет конечная температура замораживания: для закаленных растений различных по устойчивости видов винограда критическая температура, после которой быстрый обогрев приводит к гибели тканей, находится в интервале —15—17°, в то время как полная выживаемость у тех же сортов после—23 —30° соответственно обеспечивается при оттаивании со скоростью 6—12°/min. Быстрое оттаивание после слабых морозов (—10,—13°) уже не оказывает вреда. Вероятно, обезвоживание до —15° не опасно для тканей винограда (Погосян, Красавцев, 1970).
Исследованиями установлено, что при быстром оттаивании одной из причин гибели клеток винограда, особенно сильно обезвоженных внеклеточным льдом, является очень высокая скорость обогрева тканей (от 50 до 100° в минуту) в температурных интервалах от —20 до —5° и от —5 до 0°.
Высокая выживаемость клеток проявляется в тех случаях, когда при одном из указанных интервалов обеспечивается низкая скорость оттаивания тканей— в пределах 2—8°/min. Повреждение виноградного растения при быстром оттаивании является результатом сильного притока талой воды в клетку из межклеточного пространства при температурном градиенте, близком точке замерзания (Погосян, Сакаи, 1972, 1972а).
Устойчивость лозы в процессе оттаивания достигается ступенчатым всасыванием талой воды в клетки и медленным восстановлением нормального его состояния. Оптимальным условием для впитывания талой воды и частичного восстановления структуры клетки, обеспечивающим последующее безвредное быстрое оттаивание, является температура в диапазоне —5—7°.
Следовательно, можно полагать, что в природных условиях, где оттаивание в основном происходит значительно медленнее (максимум от 0,8 до 1,0° в минуту), гибель тканей лозы при этом, за редким исключением, не имеет места: она происходит в основном при критических для винограда температурах.
Таким образом, механизм приспособления тканей винограда к быстрому оттаиванию сходен с древесными с той лишь разницей, что у винограда уровень протекающих приспособительных процессов несколько смещен, что в значительной степени связано со свойствами протопласта.
Полученные результаты опытов, в частности по выживаемости тканей лозы при различных скоростях замораживания и оттаивания, могут быть подтверждены при изучении морозоустойчивости виноградного растения с точки зрения его фитоклимата.
Переходя к рассмотрению вопроса по существу, отметим, что для исследований мы использовали 8-точечный чувствительный самописец с термопарами диаметром 0,2 mm, расположенный в различных органах и тканях лозы по всей высоте куста.
Исследования фитоклимата, колебаний температур в различных частях и тканях виноградной лозы (с точностью определения ±0,2°), как и ее морозоустойчивости показали, что ткани виноградного куста даже в морозную зиму (средняя температура января —3, —5°), подвержены большим ежедневным колебаниям температуры. При этом флоэма южной стороны штамба (на высоте 10 cm выше снежного покрова) подвергается наибольшим суточным колебаниям температуры, кратковременный перепад (25—50 min) которых в отдельные солнечные дни может достигнуть 35° (от —17 до +18°). Сравнительно слабым (16—20°) колебаниям температуры подвергаются ткани почек и однолетнего побега. В отличие от надземной части растения южная сторона штамба (флоэма) на глубине 20 cm от поверхности снега в течение морозной зимы (2—2,5 месяца) находится в замороженном состоянии в диапазоне температур от —2 до —6°, в результате чего блокируется передвижение воды из корней в надземную часть куста, хотя и последняя подвергается ежедневному солнечному обогреву (Pogosian, Sakai, 1969). Аналогичное явление наблюдается и у ряда древесных растений (Sakai, 1968). В холодные зимы (средняя температура воздуха —3—5°) ежедневные кратковременные колебания температур в тканях надземной части виноградного растения не вызывают ослабления морозоустойчивости; в тканях происходит только частичное таяние льда, образовавшегося при второй фазе закаливания, поэтому последующее понижение температуры (со скоростью 0,2—0,6° в минуту) легко восстанавливает первоначальное состояние, обеспечивая относительно стабильную устойчивость тканей лозы к морозам в зимние месяцы (Погосян, Сакаи, 1972).
В зимы с длительными оттепелями (не менее 5 дней с максимальной дневной температурой до +10 + 15°) и резким похолоданием морозостойкость винограда значительно слабеет, поскольку состояние второй фазы в целом утрачивается и эффект первой фазы закаливания частично ослабевает. В данном периоде критическая температура для лозы заметно повышается, и поэтому сравнительно быстрое понижение температуры в ночное время уже проявляет отрицательный эффект. Не имеется достаточного времени для: прохождения процессов, обеспечивающих закаленное состояние тканей.
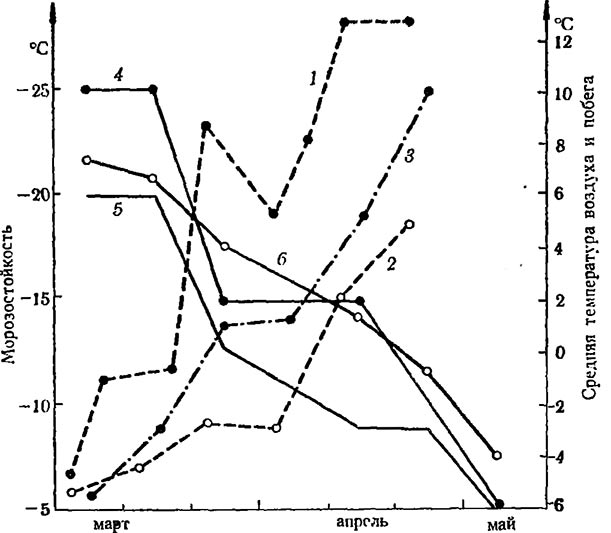
Рис. 1. Колебание температуры у основания штамба и понижение морозостойкости однолетнего побега
1,2 — максимальная и минимальная температура флоэмы штамба (юг) на глубине 20 cm ниже поверхности снега (до 19/III) и на поверхности (после 20/III); 3— средняя температура воздуха; 4— замещающая почка; 5 — главная почка; 6 — флоэма
Мы склонны утверждать, что повреждение или гибель почек и тканей лозы, в основном, наступает, когда отрицательная температура в них снижается до критической температуры, соответствующей физиологическому состоянию данного растения или отдельных его органов.
Морозостойкость лозы резко падает, когда колебания температуры затрагивают замороженную часть штамба (10—12 cm ниже поверхности снега) и поступление воды из почвы в надземную часть куста, подвергающуюся уже в этот период более длительному обогреву, становится возможным (рис. 1). Однако пока недостаточно выяснено, что является наиболее ответственным в понижении морозостойкости винограда в конце зимы и в ранневесенний период — начало поступления воды или продолжительность действия температур в пределах 10—12°.