Известно, что, несмотря на широчайшее разнообразие потребляемых и образуемых растением соединений, метаболические системы отличаются поразительной простотой и упорядоченностью. Макромолекулы различной химической природы, разрушаясь, образуют наиболее важные продукты центральных путей обмена в сопровождении биосинтеза макроэргов и сопряжении с процессами трансформации химической энергии. Рассмотрим с этих позиций вопрос о сопряженности процессов углеводного и липидного обмена в побегах в природных условиях зимовки в связи с морозоустойчивостью сорта. Сопоставляя характер кривых содержания моно-, диолигосахаридов, крахмала, сырого жира, фосфолипидов и галактолипидов в одних и тех же материалах, Дограмаджян и Марутяи (1972), Дограмаджян с сотр. (1969), Марутян и Абаджян (1976) не утверждают существование непосредственной прямой или косвенной связи между ними. Известно, что пути анаболизма и катаболизма даже через центральные пути обмена в деталях никогда не повторяют друг друга вследствие компартментации ферментов, субстратов и кофакторов.
Сопоставление кривых на рис. 1 показывает, что крахмал и сырой жир, с одной стороны, а также крахмал и сахароза — с другой, проявляют противоположное количественное изменение в течение всего осенне-зимнего периода у обоих сортов, тогда как сахароза и моносахариды — только в период минусовых температур. Наряду с такой общностью имеются и сортовые различия.
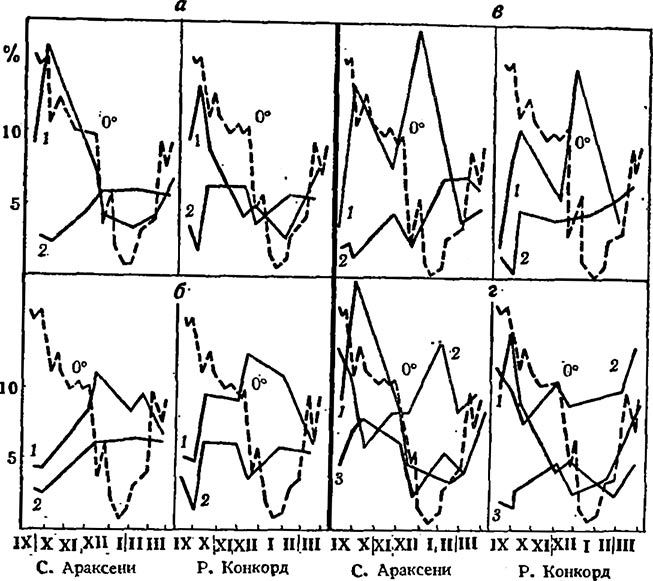
Рис.1. Сравнительная динамика содержания некоторых метаболитов в побегах винограда у сортов разной морозоустойчивости в период осенне-зимнего покоя
а — крахмал (1) и сахароза (2);б — моносахариды (1) и сахароза (2); в — галактолипиды (7) и стахиоза (2); г — крахмал (1), свободные жиры (2) и фосфолипиды (3)
У морозоустойчивых сортов в течение всего периода покоя обратная зависимость между кривыми проявляется между крахмалом и фосфолипидами, а у неустойчивых — между стахиозой и галактолипидами.
Накопление сырого жира на фоне убыли крахмала говорит об интенсивной анаэробной диссимиляции углеводов, обеспечивающих процесс биосинтеза липидов нужными метаболитами. Об анаэробиозе свидетельствует также накопление спирта в побегах винограда зимой (Рouget, 1963). Образование фосфолипидов, сопровождающих процесс катаболизма крахмала у морозоустойчивых сортов, можно рассматривать как один из примеров ступенчатого освобождения и новой аккумуляции химической энергии в условиях низких температур. Сохранение постоянства процессов распада и биосинтеза фосфолипидов как основного компонента мембран всех органелл клетки можно считать одним из важнейших факторов формирования морозоустойчивости виноградной лозы.
Энергетическое превосходство метаболических реакций морозоустойчивых сортов доказывается (табл. 3 и 4) комплексом признаков: высоким уровнем содержания нуклеотидов, белков, органических кислот, оксидоредуктаз при воздействии критических морозов.
Таблица 3
Содержание азотистых веществ, органических кислот и нуклеотидов в побегах винограда при воздействии —24° в природных условиях
| Показатель | Морозоустойчивый сорт, М±m | Неморозоустойчивый сорт, М±m |
Общий азот, mg/g | 6,28 ±0,34 | 5,64 ±0,25 |
Белковый азот, mg/g | 3,72±0,12 | 1,67±0,11 |
Небелковый азот, mg/g | 2,56±0,09 | 3,97±0,16 |
Амидный азот, mg/g | 0,16-+0,005 | 0,37 ±0,02 |
Аммиачный азот, mg/g | 0,20±0,013 | 0,04±0,001 |
Сумма свободных аминокислот, mg/g | 4,6±0,28 | 16,8±0,83 |
Отношение белкового азота к небелковому | 1,45 | 0,42 |
Общее количество органических кислот, mg/g | 33,65 ±1,31 | 27,50±1,8 |
Ди- и трикарбоновые кислоты, mg/g | 18,45 ±0,93 | 8,17 ±0,64 |
Винная кислота, mg/g | 13,35±0,81 | 7,91 ±0,40 |
Лимонная кислота, mg/g | 2,80±0,13 | следы |
АТФ, pg/g | 5,60±0,21 | 4,16±0,19 |
АДФ, pg/g | 5,03 ±0,14 | 3,22±0,15 |
АМФ, pg/g | 1,86±0,06 | 1,40 ±0,06 |
Сумма нуклеотидов, pg/g | 12,49 ±0,50 | 8,78+0,35 |
Таблица 4
Активность ферментов в побегах винограда при воздействии —24° в природных условиях
Показатель |
| Морозоустойчивый сорт, М±m | Неморозоустойчивый сорт, М±m |
Протеазы | mg азота за 48 часов | 55,86±4,1 | 98,96 ±6,8 |
Амилазный комплекс | mg глюкозы за 48 часов на 1 g навески | 67,0 ±2,4 | 44,4 ±2,9 |
Каталаза | cm3 0,1 n Н2O2 за 6 минут на 1 g |
| |
| навески | 20,8 ±1,3 | 6,9 ±0,56 |
Пероксидаза | mg пурпурогаллина за 10 минут на 1 g навески | 6,25 ±0,28 | 0,92 ±0,07 |
Полифенолоксидаза | mg пурпурогаллина за 10 минут на 1 g навески | 1,50±0,10 | 2,00±0,13 |
Присутствие малоната только в побегах морозоустойчивых сортов, сопровождающееся высоким содержанием магния и серы, динамичностью АТФ и SH-группы, белков, оксидоредуктаз, возможно, указывает на более эффективное использование химической энергии у них в зимний период.