Выведение и улучшение сортов винограда на основе применения методов мутагенеза, полиплоидии и клоновой селекции
Из курса генетики (науки о наследственности и изменчивости организмов), известно, что организмы построены из клеток, в которых имеются ядра, содержащие хромосомы (рис. 82). Хромосомы состоят их генов - основных единиц передающих наследственную информацию. Ген же по сути представляет собой участок ДНК (дизоксинуклеиновая кислота), в структуре которой зашифрована программа наследственности.
При размножении и развитии организмов происходит деление половых и соматических клеток. При делении клеток делятся и ядра, а дочерние клетки получают по новенькому ядру, полностью повторяющему структуру старого. Если в старом ядре было сорок или двадцать хромосом, то столько же сохраняется и в ядрах дочерних клеток. Перед делением хромосомы удваиваются и, в процессе деления, удаляются друг от друга, отходя к противоположным полюсам клетки.
Способ деления клеток, при котором вновь возникающие клетки сохраняют исходное число хромосом носит название м и т о з. Каждая хромосома содержит ДНК и ряд различных белков. Нуклеиновых кислот в хромосомах соматических клеток содержится очень мало, около шести миллиардных долей миллиграмма ДНК. Однако при передаче такого количества ДНК дочерним клеткам вполне достаточно, чтобы сохранилась наследственная информация для будущих поколений, т.к. ДНК занимается “организацией и руководством” строительства будущей клетки.
Кроме митотического клеточного деления существует еще процесс образования половых клеток - гамет, который носит название мейоз.
Половые клетки-гаметы (яйцеклетки или сперматозоиды) имеют одинарный набор хромосом. При их образовании происходит два последовательных деления следующих одно за другим. Во вновь образовавшиеся клетки попадает вдвое меньше хромосом, чем их было в отцовской, где каждая хромосома была представлена двумя копиями или гомологами. Мейоз разгоняет эти копии по гаметам, оставляя в яйце или сперматозоиде по одному набору хромасом. В процессе оплодотворения гаметы сливаются и получается зигота-оплодотворенное яйцо уже с нормальным двойным набором хромосом.
Клетки с одинарным набором хромосом называются гаплоидными. Сюда относятся гаметы высших растений и животных, а также клетки низших растений и бактерий в обычном состоянии.
При слиянии половых клеток возникает нормальный двойной набор хромосом, который получил название диплоида. Диплоидными обычно являются соматические клетки высших растений и животных. Хромосомы диплоидных клеток в два раза тяжелее хромосом гаплоидов.
Кроме гаплоидов и диплоидов имеются еще полиплоидные клетки с нескольким набором хромосом (тетраплоиды - 4 набора хромосом и ДНК в них в 4 раза тяжелее, чем у гаплоидных).
Взвешивание ДНК у гаплоидных и полиплоидных клеток еще раз подтверждает, что главные факторы наследственности находятся в хромосомах, что ДНК содержит основную наследственную информацию клетки.
До деления (митоз или мейоз) клетка находится в состоянии относительного покоя, обмен информацией идет между ядром и цитоплазмой. Клетка как бы преодолевает время и сохраняет пространство. Когда же наступает деление, то клетка быстро захватывает новое пространство за короткий промежуток времени. Митоз или мейоз обозначают как бы информационный взрыв в пределах клетки. Что же является сигналом к нему? Это пока не разгадано. Некоторые считают, что сигналом может служить соотношение объемов ядра и цитоплазмы. Рост ядра идет медленнее, чем рост цитоплазмы, поэтому внутри клетки создается авральное положение - много строительных материалов в цитоплазме, а места не хватает и расхода нет, что затрудняет обмен веществ. Вот в эти моменты ядро начинает делиться, чтобы увеличить командную поверхность (2 ядра) и уменьшить цитоплазматический объем. Считают, что клетка сама улавливает приближение авральной ситуации и подает сигналы к делению.
Самым удивительным итогом деления клеток является создание идентичной копии. Клетка из клетки, организм из организма, дети похожи на родителей, от кошки не рождаются волки - таковы внешние проявления законов наследственности. Это стало хорошо понятным и объяснимым только при создании хромосомной теории, начиная с открытия законов Грегором Менделем (1866) и установления в наши дни роли генов, ДНК и РНК, а также других химических носителей наследственности.
По Менделю каждому чистому наследственному признаку соответствует один ген. Если в одной хромосоме находятся два гена, то они обычно передаются потомству вместе. Это сцепленные гены. Гены, создающие разные признаки, находятся в разных хромосомах и передаются независимо друг от друга. Это подтверждается способностью хромосом разрываться и обмениваться частями (кроссинговер). При этом материнские и отцовские гены меняются местами в хромосомах.
Все сложные признаки организмов создаются не одним, а множеством генов.
Гены способны мутировать, т.е. резко изменять качество передаваемой наследственной информации. Мутации генов совершаются всегда резко, скачкообразно. При этом гены, обретя новую форму, сохраняют и передают ее наследникам с такой же автоматической непреклонностью с какой они передавали старую информацию. Новый ген также устойчив и постоянен, как и любой другой. Поэтому мутация не связывается с обычными наследственными комбинациями генов. При мутациях организм изменяется коренным образом по сравнению с обычной неследственной изменчивостью. Например, можно сравнить наследственную изменчивость с погодой, а мутацию с землетрясением, когда складки земли деформируются.
Считают, что мутация вызывает нарушение структуры ДНК поэтому меняется характер передающей информации. Это примерно то же самое, что менять порядок букв в словах или слов во фразах. Не выяснено только происходят ли перемены в самом составе ДНК.
Человеку издавна было известно большое число случаев, когда среди чистых сортов растений и пород животных появлялись отдельные формы, обладающие некоторыми сильно отличающимся признаками, которые в дальнейшем сохранялись в потомстве.
Так, еще в 1590 г. в одном из садов Гейдельберга отметили появление ланцетовидной формы Chelidonium mayis, а в конце 17 века в новой Англии появилась овца с очень короткими ногами, которая стала родоначальницей породы анконских овец. В Швеции появился ячмень с ослепительно зеленой окраской. В природе существует большое количество альбиносов.
О таких изменениях знал еще Чарлз Дарвин. Он их называл “единичными изменениями” и “спортами”.
Впервые мысль о резком скачкообразном изменении живого высказал крупный французский ученый Этьен Жофруа Сент-Илен (1772-1844). Позже об этом отмечал академик Коржинский (1899), затем голландский ботаник Гуго-де-Фриз, который предложил термин “мутация” в применении к биологии. Он считается отцом мутационной теории.
Все мутации, возникающие в естественной природе, естественных условиях называют спонтанными.
По закону гомологических рядов академика Н. И. Вавилова спонтанные мутации возникают у всех видов растений, и особенно на границе ареала их распространения. Однако они возникают крайне редко. По данным Хенкена у картофеля обнаружена одна мутация на 200 тыс. растений. Частота их возникновения зависит и от вида растений. По данным Бауэра (1922) у растения львиный зев на 1000 растений приходится два мутанта, причем большая часть мутаций представляет собой небольшие количественные изменения, так называемые малые мутации. В результате многократного возникновения малых мутаций возникает изменения и эволюционного характера.
Полезных для человека мутаций возникает значительно меньше. Поэтому человек начал искать методы и приемы, с помощью которых можно вызывать искусственное появление мутаций. Такие мутации, в отличии от самопроизвольных-спонтанных, называют искусственными, экспериментальными, индуцированными.
Индуцированные мутации получают при использовании ионизирующих излучений (гамма-облучение, ультрафиолетовые лучи, рентгеновские лучи, ИКСС - импульсный концентрированный солнечный свет и другие виды лучистой энергии) - это так называемый радиационный мутагенез.
Кроме этого мутации получают путем обработки разными химическими веществами - химический мутагенез.
Искусственный мутагенез применяют в следующих случаях:
- При межвидовых и более отдаленных скрещиваниях для получения сортов с комплексной устойчивостью к болезням и вредителям, когда гибриды первого поколения, как правило, наследуют признаки дикого качества одного из родителей.
- При получении новых свойств и качеств полезных для человека, которые раннее отсутствовали.
- Для нарушения тесного сцепления между полезными и нежелательными признаками у исходных сортов и гибридных сеянцев, получение мутантов и использование их для последующих скрещиваний.
- Для расширения границ изменчивости в гибридных популяциях сортов и создание генофонда мутантов.
- Для преодоления нескрещиваемости и стерильности гибридов при межвидовых и межродовых скрещиваниях.
- Для подавления диких признаков у гибридов первого поколения, путем облучения пыльцы дикого вида или сорта.
В настоящее время известны и применяются следующие химические мутагены: ЭИ - этилен имин; ДМС - диметил сульфат; НЭМ - нитрозо- этилмочевина; колхицин .
Химические мутагены применяют пользуясь следующими способами:
- вымачивание семенного или черенкового материала в водном растворе или суспензии мутагенов;
- внедрение раствора или суспензии мутагена в семена методом вакуумфильтрации;
- внедрение раствора или суспензии мутагена в семена методом ультразвуковой обработки;
- введение раствора или суспензии мутагена в растение или отдельные побеги методом инъекции или нагнетания.
Обработка семян химическими мутагенами проводится или без дополнительной подготовки, или после подготовки семян, а именно:
- вымачивание воздушно-сухих семян в растворе мутагена без какой- либо предварительной подготовки;
- обработка мутагенами семян после их вымочки в воде в течение трех суток до состояния набухания;
- обработка семян мутагенами после их стратификации в песке при температуре 20-25°С.
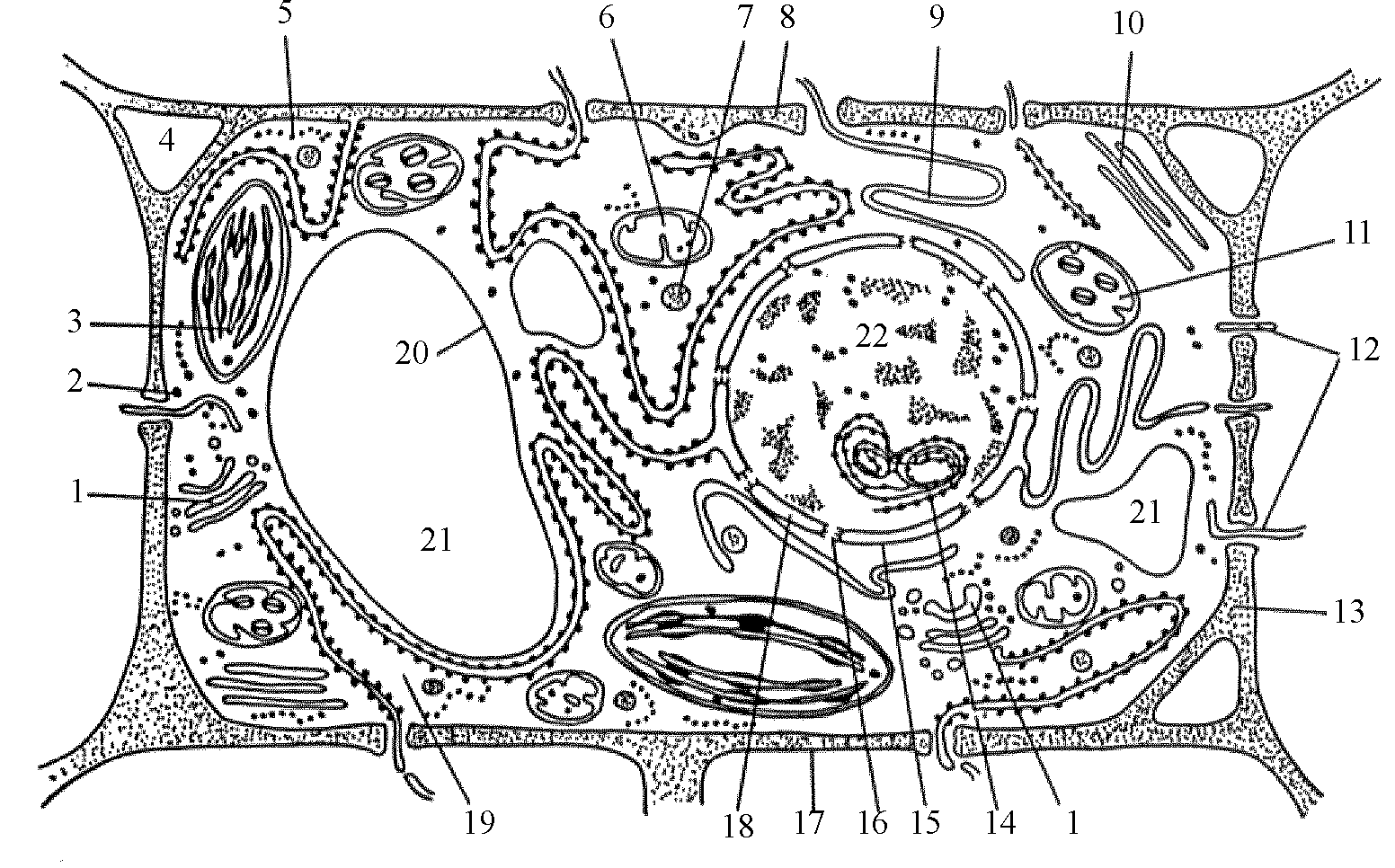
Рис. 82. Обобщенная схема растительной клетки:
(1 — аппарат Гольджи; 2 — свободно расположенные рибосомы; 3 — хлоропласты; 4 — межклеточные пространства; 5 — полирибосомы; 6 — митохондрии; 7 — мезосомы; 8 — гранулированная эндоплазматическая сеть; 9 — гладкая эндоплазматическая сеть; 10 — микротрубочки; 11 — пластиды; 12 — плазмодесмы, проходящие сквозь оболочку; 13 — клеточная стенка; 14 — ядрышко; 15, 18 — ядерная оболочка; 16 — поры в ядерной оболочке; 17 — плазмолема; 19 — талоплазма; 20 — тонопласт; 21 — вакуоли; 22 — ядро).
Таблица 10
Примерные концентрации мутагенов при обработке воздушносухих семян (срок вымочки 24 часа)
При обработке набухших или надтреснутых семян винограда дозы всех трех мутагенов можно увеличить для первых двух до 0,5-1% и для третьего до 0,1%.
Обработка вегетирующих вегетативных и генеративных органов химическими мутагенами проводится методом принудительной подачи мутагена под давлением 0,2-0,3 атмосферы, которое создается с помощью насоса Камовского. Раствор подается по трубкам (резиновым или пластмассовым) к побегам, которые отделяются от кустов и на их нижний конец одевается подводящая трубка, по которой нагнетается мутаген в побеги, в количестве до 15 мл.
Обработанные мутагенами побеги затем прививаются зеленой прививкой на материнские кусты. В следующем году с обработанных глазков получают урожай или, если побеги имели соцветия, оценку урожая возможно сделать и в год прививки.
Следовательно, обработка мутагенами вегетирующих побегов ускоряет селекционный процесс по сравнению с обработкой мутагенами вызревших черенков. Концентрации растворов НЭМ - 0,001%, ДМС - 0,1%.
Радиационный мутагенез или обработка семян, соцветий, вегетативных органов гамма-лучами (Со60). Источником облучения служит гамма-установка марки ГУТ-Со400, выпускаемая промышленностью. Мощность дозы от 1 до 5 р/сек. В качестве источника гамма-лучей можно использовать изотоп цезий - 137, а также источники нейтронов, колонки атомных реакторов, ускорители зараженных частиц, рентгеновские лучи, ИКСС - импульсный, концентрированный солнечный свет и другие виды лучистой энергии.
Продолжительность обработки семян зависит от их состояния (воздушно-сухие, набухшие, надтреснутые) и от мощности заряда источника. Рабочими дозами при облучении семян винограда с целью получения мутаций следует считать 4,5-8 тыс. рентген. При этом выживает 10-50% сеянцев. Летальной дозой для семян винограда любого происхождения является 10-12 тысяч рентген.
Облучение соцветий винограда производят вместе с зелеными побегами, на которых они расположены на стационарных установках. После облучения побеги нарезают на одноглазковые черенки, которые вместе с соцветиями прививают в зеленом виде на кусты подвоя методом простой копулировки. В этом же году соцветия образуют грозди. Осенью из созревших ягод извлекают семена, из которых затем выращивают сеянцы. После вступления сеянцев в пору плодоношения ведется отбор на выявление мутагенных форм с хозяйственно-ценными признаками.
Облучение соцветий проводится в два срока: за 1,5-2 недели до цветения и во время цветения. В первом случае облучение действует на материнские клетки будущих гамет, во втором на клетки зигот перед вступлением их в метотическое деление.
Доза облучения в первый срок 2,5 тыс рентген, во второй - 2 тыс рентген при мощности установки гамма облучения 500 р/мин.
Облучение вегетативных органов - растущих зеленых побегов винограда. Когда побеги достигают длины 10-12 междоузлий их срезают с материнских кустов помещают в полиэтиленовые мешочки с увлажненной марлью и подвергают гамма-облучению на стационарных установках. После этого побеги нарезают на одноглазковые черенки. Черенки 4-5 узлов затем прививают методом копулировки, при этом на узлах оставляют или только зимующий глазок или только пасынок. Используются следующие дозы облучения:
- при мощности излучения установки 250 р/мин - 500-7000 рентген;
- при мощности излучения установки 500 р/мин - 2500-5000 рентген.
В следующем году, побеги выросшие из гамма-облученных формирующихся глазков или пасынков способны плодоносить. С этого года можно вести отбор кустов по количеству и качеству урожая.
Облучению можно подвергать и одноглазковые одревесневшие черенки используемые при настольной прививке. Рабочими дозами в этих случаях являются 3000-5000 р. а летальной дозой - 12000 рентген.
Облучение пыльцы. Пыльцу собирают обычным способом и раскладывают в пакетики для обработки гамма-лучами разными дозами на установках ГУТ Со 60-400 или ГУБЭ - 4000 и др. Дозы облучения - 10, 15, 20, 25 и 30 кр. Для сохранения жизнеспособности пыльцы срок от сбора до опыления должен быть не более 5-7 дней.
Полиплоидия в селекции винограда как и на многих других растениях может оказать содействие в развитии желаемых признаков у новых сортов винограда при гибридизации, в связи с более богатой генетической информацией полиплоидных форм.
Полиплоидные растения зачастую имеют большие размеры клеток у отдельных органов, измененные морфологические и биологические признаки, увеличенный урожай. Известно, что большинство сортов винограда Vitis vinifera имеет 2n=38, а у видов подрода Мускадиния 2n-40.
Исследования ряда ученых показали, что среди представителей вида V. Vinifera имеются формы с 2n=76 хромосом. Это тетраплоиды.
Тетраплоидные формы отличаются от диплоидных особенностями строения и развития вегетативных и генеративных органов, в частности укороченными междоузлиями, более толстыми побегами, крупными кожистыми, темнозелеными листьями с хорошо обозначенными семью жилками, более крупными соцветиями, цветками, пыльцевыми зернами, гроздями, ягодами и семенами.
Истинными тетраплоидами являются сорта Шасла Грокуляр белая и розовая. Имеются тетраплоиды среди выделенных крупноягодных клонов сортов Португизер, Рислинг рейнский, Рислинг итальянский, Шабаш, Кишмиш белый, Кишмиш черный, Мускат Александрийский и др. Не исключено, что такие же формы имеются среди других сортов вида V.Vinifera.
Поэтому необходимо вести селекцию винограда и на полиплоидном уровне. Для этого собирается коллекция уже известных крупноягодных форм, на которых нужно проводить цитологический контроль, а затем включать в гибридизацию, обработку мутагенами и другими существующими методами.
Так, ИВиВ “Магарач” выделены тетраплоидные сорта, которые были использованы в гибридизации с диплоидными сортами. В результате таких скрещиваний была получена гибридная форма (2n=57) - триплоид, названный сортом Поливитис Магарача. Этот сорт получен в результате скрещивания сорта Катта Курган (2n=38)χШасла грокуляр белая (2n=76). Новый сорт обладает комплексом ценных хозяйственных признаков, а именно большой размер грозди (537-564 г), повышенное содержание сахаров, более ранние сроки созревания. Поливитис Магарача отличается также сильным ростом куста, мощными побегами и высокой урожайностью. На Южном Берегу Крыма при площади питания 3-3,5*2 м, мощных штамбовых формировках и короткой обрезке был получен в среднем за 2 года урожай по 9,9 кг с куста при средней массе грозди 750 г, средней массе ягоды 6 г, сахаристости 17% и кислотности 8,5 г/л. В расчете на 1 га урожайность составила 142,8 ц.
Для улучшения сортов и выявления новых форм среди стародавних сортов винограда в практике виноградарства широко используется клоновая селекция.
С давних времен в садоводстве и виноградарстве известны случаи наследственной изменчивости отдельных признаков и свойств многолетних культур.
Древнеримские ученые Колумелла, Паладий и другие наблюдали подобные факты у винограда и рекомендовали использовать их в практических целях. Колумелла писал, что при размножении необходимо отбирать потомство лишь самых плодовитых лоз, отчеренковывая наиболее урожайные их части. Он утверждал также, что медленный ход процесса не должен никого отпугивать. Проводить клоновую селекцию рекомендовали известные французские виноградари Гюйо, Казенава, Каррьер и др. В Германии этими же вопросами занимался О. Сарториус, в Швейцарии - Мюллер Тургау, в Калифорнии - Ф. Биолетти. В СССР горячими сторонниками клоновой селекции и ее пионерами можно считать Мержаниана, Мельника, Папонова, Цебрия и др. Особенно настойчиво советовал применять отбор лучших растений, лоз и черенков И. В. Мичурин: “Тщательным отбором черенков, повторением отводки лучших частей лозы, сравнительно короткой обрезкой и посадкой на лучшую почву следует способствовать развитию лучших качеств”.
Наличие почковой изменчивости у многолетних растений в настоящее время общепризнанный факт, поэтому задачи повышения урожайности и улучшения качества продукции можно решать не только путем выведения сортов винограда на основе половой гибридизации, но и в результате использования вегетативной изменчивости известных стародавних или новых сортов, вызванной естественными или искусственными факторами.
Различают два типа вегетативной изменчивости:
- наследственная изменчивость, которая проявляется при вегетативном размножении винограда в виде почковых мутаций, которые возникают на базе колоний мутантных клеток. Колонии клеток могут произойти от одной соматической клетки - носительницы нелетальной мутации;
- ненаследственная вегетативная изменчивость. Она очень разнообразна и представлена обычными и длительными модификациями- флюктуациями.
Существуют также вариации вегетативной изменчивости: морфологическая, физиологическая, биохимическая, кариотипическая (изменение числа хромосом).
Клоны с желаемой изменчивостью нужно отбирать.
Они могут быть положительные (полезные) и отрицательные (нежелательные).
Известны случаи, когда новые ценные сорта через некоторое время после введения их в производство оказываются затерянными, в результате сильного засорения низкокачественными и малоурожайными клонами. Это особенно сильно проявляется если для размножения используются еще неплодоносящие и неподвергшиеся отбору растения.
Даже на самых высококачественных плантациях со временем появляются отрицательные кусты и если не проводить работу по клоновой селекции, их число будет все время увеличиваться.
Клоновая селекция это один из действенных широко доступных этой селекции. Так, в Германии за последние 40 лет благодаря клоновой селекции продуктивность виноградных насаждений возросла почти в 4 раза. Результаты работ по клоновой селекции в НИИВиВ в Братиславе показали, что у выделенных клонов многих сортов винограда урожайность была выше в 2-3 раза по сравнению с неселекционными кустами. При этом кислотность и сахаристость ягод существенно не изменялись.
В Республике Молдова также получено много новых клонов среди сортов Пино черный, Траминер, Шардоне и др., превышающих сорт по урожайности в 2-3 раза.
Подсчеты ученых показывают, что урожайность отдельных сортов с помощью клоновой селекции можно повысить в среднем на 20-30%.
Под клоном понимают вегетативно размноженное потомство одного куста или отдельного побега, отличающегося от исходного сорта новыми стойко закрепившимися хозяйственно-ценными свойствами, наследуемыми при вегетативном способе размножения.
Для отбора практическое значение имеют лишь хорошо выраженные почковые мутации и длительные модификации ценных признаков и свойств. Причем превосходство отобранных клонов над исходным сортом по одному или комплексу признаков должно быть подтверждено на всех этапах селекции статистическими методами обработки экспериментальных материалов.
В зависимости от целей работы клоновая селекция может быть массовой или индивидуальной.
Массовая селекция (отбор) проводится с целью поддержания сорта в оптимальной продуктивности и фенотипической выраженности. Сорт при этом улучшается постепенно.
Массовую селекцию в зависимости от качества виноградных насаждений проводят двумя способами:
- По отрицательным признакам, когда на винограднике выделяют малоурожайные, осыпающиеся, горошащиеся, больные и другие малоценные кусты основного сорта и примеси других сортов.
- По положительным признакам, когда отбирают только кусты основного сорта с хорошими показателями по урожайности, силе роста и качеству урожая. Наиболее экономически выгодно отмечать этикетками и записями в полевых журналах кусты с теми признаками, которых на участке меньше.
Индивидуальную клоновую селекцию проводят путем отбора и размножения самых лучших по тем или иным признакам кустов с целью повышения хозяйственной ценности районированных сортов.
Методика проведения индивидуальной клоновой селекции предусматривает работу как минимум в 4 этапа:
- Выявление форм вегетативной изменчивости и определение направления отбора, а также выбор участков.
Наиболее распространенным направлением клоновой селекции является отбор на повышение урожайности в сочетании с сохранением или улучшением качества ягод у сорта. Для этой цели селекция ведется по следующим основным показателям:
- высокая и постоянная урожайность при хорошем качестве урожая;
- хороший рост кустов сочетающейся с необходимыми показателями урожайности и качества урожая;
- отсутствие инфекционных заболеваний.
Очень важным также является отбор на раннеспелость, крупноплодность, высокую сахаристость и т.д.
Работу начинают с детального ознакомления с виноградниками тех сортов, которые намечены для отбора. В процессе ознакомления отбирают участки, на которых имеются желаемые формы и изменения и останавливаются на конкретном участке, который должен быть по возрасту от 7 до 20 лет.
Обследование виноградников проводят перед сбором урожая. Выявленные кусты с полезными формами изменчивости (урожайность, величина и форма грозди и ягоды, срок созревания и т.д.) фиксируются глазомерно и определяется примерный их удельный вес. Состояние кустов должно быть хорошее.
- Отбор маточных кустов их оценка и размножение.
После установления направления отбора на участке отбирают кусты, отличающиеся повышенной урожайностью в сочетании с хорошим ростом и нормально развитыми ягодами, нормальными или более крупными гроздями с нормальной окраской и равномерным созреванием.
Отобранные кусты отмечают этикетками и каждому из них присваивают номер, который сохраняется за ним и его потомством в период дальнейшего испытания и размножения.
При сборе урожая с каждого выделенного куста срезаются грозди в отдельности и пересчитываются, затем взвешиваются и результаты записываются в специальный журнал учетов и наблюдений, проводимых на клоновом участке. Параллельно проводят учеты и наблюдения и на 10-15 кустах - эталонах, которые отбираются методом сетки. Данные по кустам эталонам служат в качестве контрольных для сравнения.
Кроме урожая учитывают число оставленных при обрезке глазков, число развившихся побегов, из них плодоносных, и число соцветий. На основании этих данных делают расчеты плодоносных побегов в %, коэффициентов плодоносности и плодоношения, плодоносности глазка, побега в г и др.
Показатели урожайности и сахаристости отобранных кустов должны быть на уровне или выше контрольных. Если эти показатели ниже, кусты исключают из дальнейшей работы. Выбраковывают также кусты имеющие особо опасные вирусные заболевания (короткоузлие, инфекционный хлороз, скручивание листьев и др.).
Оценку кустов, отобранных в первый год, проводят по той же схеме и на второй год. Кусты сохранившие ценные свойства выделяют в качестве маточных для клоноиспытания.
Осенью второго года с отобранных маточных кустов (с каждого в отдельности) заготавливают черенки и связывают в пучки, на которые навешивают этикетки с названием сорта и селекционным номером куста.
Иногда для ускорении селекционного процесса заготовку черенков ведут с первого же года, при этом учитывают, что саженцы, выращенные из забракованных во втором году кустов, будут в дальнейшем исключены. За контроль принимаются саженцы, выращенные из общей партии черенков, заготовленных на участке со здоровых кустов.
В зависимости от зоны виноградарства саженцы могут быть корнесобственные или привитые.
- Первичное клоноиспытание.
Саженцы, выросшие из черенков маточных кустов в количестве не менее 10 шт, высаживаются на участок первичного клоноиспытания. Посадка клонов ведется последовательно номер за номером в стандартные ряды длиной 100 метров по 60 кустов или 6 клоносмесей. Допускается посадка каждого клона по 20 и более кустов. Посадка проводится без повторений.
На каждые 4 ряда клонов высаживают 20 кустов контрольной партии (популяции сорта). Наиболее ценные номера клонов усиленно размножают. Агротехника на участках клоноиспытания обычная, согласно агроуказаний.
Учеты и наблюдения на клоновых посадках начинают с 4 года после посадки и ведут их в течение 3 последних лет нормального плодоношения.
Учеты и наблюдения проводят по методике первичного сортоизучения, применяемой в ампелографических коллекциях.
По результатам трехлетней оценки элитные клоны выделяют для конкурсного испытания.
- Конкурсное испытание проводят по методике государственного испытания и завершают его выделением суперэлиты.
Как и на предшествующем этапе клоноиспытания для ускорения работ рекомендуется начинать размножение лучших клонов заблаговременно, не дожидаясь окончания клоноиспытания.
Необходимо знать, что внешний осмотр не всегда обеспечивает получение безвирусного посадочного материала. Поэтому, выделенные в результате испытания клоны, должны быть проверены на зараженность вирусами на индикаторных растениях, а при выявлении заболевания проходить термотерапическую процедуру, выполняемую в специальных вирусологических лабораториях.
При окончательной оценке посадочный материал забракованных клонов изымается из размножения и передается для производственных посадок.
Если в результате первичного и конкурсного испытания выявлены особо ценные клоны, то их размножают для государственного и производственного испытания.