Как многолетнее плодовое растение виноград состоит из двух групп органов. Вегетативные органы — корень, стебель, лист, почки, усик — выполняют функцию жизнеобеспечения растения. Они поглощают и перемещают воду, питательные вещества, в них происходят важные процессы метаболизма (фотосинтез, дыхание и др.), выполняют функцию роста, вегетативного размножения (части стебля) и т. д.
Генеративные (репродуктивные) органы — соцветия, цветки, грозди, ягоды, семена — выполняют функцию полового размножения (у семенных форм). Развитие генеративных органов заканчиваются формированием съедобных ягод, используемых человеком для разных целей. Как и любое другое многолетнее плодовое растение, виноград состоит из надземной и подземной частей.
3.2.1. Корень и корневая система
Корень выполняет ряд жизненно важных функций. Он выполняет якорную функцию, закрепляя виноградное растение в почве (субстрате). Важнейшая функция корня — это поглощение из почвенной среды воды, питательных элементов, транспорт их в стебель. По проводящей системе корней также осуществляется транспорт органических веществ. В корнях винограда накапливается большое количество запасных питательных веществ и происходят сложные биохимические процессы, в результате которых вовлекаются в метаболизм поглощенные минеральные вещества, образуются фитогормоны и т. д. Корень осваивает почвенную среду, взаимодействуя с ней. При этом корень выделяет в почву некоторые продукты метаболизма, осуществляет симбиотическое взаимодействие с микоризой.
У растений, полученных из семян, — сеянцев — корень развивается из меристемы зародышевого корешка. При этом образуется главный (стержневой) корень, который направляется вертикально в глубь субстрата; очень быстро на нем развиваются корневые волоски, и они начинают выполнять функцию поглощения. По мере развития главного корня на нем закладываются корни следующего порядка ветвления и т. д. Корни следующих порядков закладываются в периферическом слое клеток центрального цилиндра — перицикла, который обладает меристематической активностью. В процессе последовательного ветвления главного корня формируется корневая система — совокупность корней разных порядков ветвления. Корневая система у сеянца имеет преимущество в развитии, а боковые корни менее развиты (рис. 4). У сеянца имеется корневая шейка — место перехода корня в стебель.

Рис. 4.
Корневая система сеянца:
1 — стержневой корень;
2 — боковые корни;
3 — корневая шейка
У саженцев, т. е. растений, полученных вегетативным путем (только так размножают культурные сорта), образуются придаточные (адвентивные) корни. На стебле отсутствуют зачатки корней.
При укоренении стеблевых частей придаточные корни образуются из клеток перицикла. Виноград обладает высокой корнеобразующей способностью. При создании соответствующих условий на укореняемых черенках образуется большое количество придаточных корней, а корневая система, формирующаяся при этом, имеет мочковатый характер. На части черенка, размещенной в субстрате, придаточные корни преимущественно образуются на узлах. Подземная часть куста, полученного вегетативным путем, состоит из подземного ствола (подземного штамба), образующегося из части черенка, помещенной в почву, и ярусов адвентивных корней. На нижнем узле подземного штамба (пятке) образуются главные, или пяточные, корни, они проникают на самую большую глубину, обеспечивают устойчивость растения к недостатку влаги. В средней части подземного штамба формируются ярусы боковых корней, они размещаются в слоях почвы, за которыми осуществляется регулярный уход. В верхней части подземного штамба непосредственно под поверхностью почвы образуются росяные (поверхностные) корни. В школке и на молодых виноградниках проводят катаровку — удаление поверхностных корней, что создает условия для лучшего развития главных и боковых корней и делает корневую систему более устойчивой при пересыхании и промерзании почвы. Особенно важен этот прием для привитой школки и привитых виноградников. Нельзя допускать образования поверхностных корней выше места прививки.
Для виноградного растения характерна мощная, развитая корневая система. Корни проникают на большую глубину, в отдельных случаях до 14 м. Боковые корни простираются на значительное расстояние в горизонтальном направлении. На последнем порядке ветвления располагаются тонкие мочки корней, которые выполняют функцию взаимодействия с почвенной средой. Строение конечной части корня не зависит от способа размножения. На самом кончике размещается корневой чехлик — конусовидное образование, состоящее из толстостенных клеток, плотно прилегающих друг к другу. Чехлик прикрывает нежные клетки апикальной меристемы, которые образуют зону роста корня (длиной около 1 мм). За зоной роста располагается зона растяжения, клетки которой, оставаясь недифференцированными, интенсивно растягиваются, длина этой зоны 3...5 мм. Быстрое деление и растяжение в зонах роста и растяжения приводит к интенсивному поглощению воды из почвы, а также проталкиванию чехлика в новые слои почвенной среды. При этом поверхностные слои клеток чехлика слущиваются, а часть апикальной меристемы, прилегающая к чехлику, формирует новые слои клеток чехлика. За зоной растяжения происходит дифференцировка клеток. Наружный слой клеток корня (эпидермис) образует выросты — корневые волоски, которые плотно прилегают к частицам почвы и резко увеличивают площадь контакта корня с почвой. Клетки эпидермиса в зоне корневых волосков — зоне поглощения — имеют целлюлозную оболочку, снаружи которой образуются выделения, улучшающие контакт с почвой. Они имеют крупные вакуоли, а цитоплазма сосредоточена вдоль стенок. Длина зоны поглощения корня достигает 2 см. По мере роста корня зона поглощения перемещается акропетально, более старые корневые волоски, расположенные дальше от кончика, отмирают, на кончике образуются новые. Продолжительность жизни корневых волосков зависит от скорости роста корня: чем быстрее растет корень, тем быстрее перемещается зона поглощения и тем меньше живут корневые волоски. За зоной поглощения располагается проводящая зона корня — здесь отсутствуют корневые волоски и теряется функция поглощения.
Процесс роста и дифференцировки клеток и тканей корня происходит по следующей схеме. Апикальная меристема расположена непосредственно под чехликом и состоит из трех слоев инициальных клеток: дерматогена, периблемы и плеромы. В результате деления, роста и дифференцировки их формируются элементы первичного строения корня: клетки дерматогена восстанавливают чехлик, а также формируют эпидермис, периблемы — первичную кору, а плеромы — центральный цилиндр. В зоне поглощения корень имеет первичное строение (рис. 5).
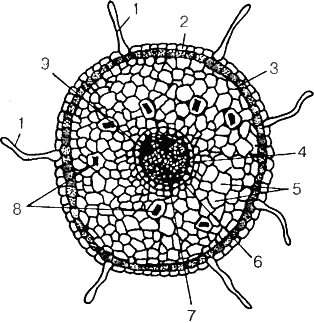
Рис. 5. Первичное анатомическое строение корня винограда в зоне поглощения: 1 — корневые волоски; 2 — эпидермис; 3 — ингеркутис; 4 — эндодерма; 5 — первичная кора; 6 — первичная флоэма; 7 — первичная ксилема; 8 — клетки, содержащие рафиды; 9 — перицикл
На поперечном срезе можно увидеть следующие его элементы: снаружи корень покрыт эпидермисом, состоящим из одного слоя клеток, образующих корневые волоски. Под ними располагается экзодерма (интеркутис), выполняющая защитную функцию. У представителей европейско-азиатского винограда экзодерма состоит из одного слоя плотно расположенных клеток, а у представителей американских видов — двух слоев. Стенки основной массы клеток интеркутиса, кроме расположенных напротив корневых волосков, опробковевают. Это обеспечивает защиту молодому корню при сохранении его взаимодействия с клетками эпидермиса и почвенной средой. Под экзодермой располагается паренхима первичной коры (мезодерма), состоящая из 10...25 слоев крупных тонкостенных клеток почти округлой формы с крупными межклетниками. Клетки коровой паренхимы выполняют функцию проведения воды и питательных элементов к центральному цилиндру, и в них накапливаются запасные вещества. За коровой паренхимой располагается эндодерма, состоящая из одного слоя тонкостенных, плотно прилегающих друг к другу клеток, выполняющих защитную и транспортную функции. Стенки части клеток эндодермы опробковевают, кроме так называемых «пропускных» клеток, расположенных напротив сосудов ксилемы и обеспечивающих поступление воды и питательных веществ в центральную часть корня.
Эпидермис, интеркутис, коровая паренхима и эндодерма составляют первичную кору корня.
Первичная кора окружает центральный цилиндр. С наружной стороны центральный цилиндр окружен несколькими слоями клеток перицикла, обладающего периодической меристематической активностью и дающего начало боковым корням следующих порядков ветвления. К центру от перицикла располагаются проводящие пучки радиального типа, включающие в себя сосуды первичной ксилемы, первичной флоэмы и паренхиму. В центре размещается слаборазвитая сердцевина.
По мере роста корня изменяется анатомическое строение и формируются элементы вторичного строения (рис. 6). Вначале появляется звездообразно-извилистое кольцо вторичной образовательной ткани — камбия. В части кольца, где камбий находится между сосудами первичной флоэмы и первичной ксилемы, он формируется из клеток основной паренхимы, а в межпучковой части — из клеток перицикла.
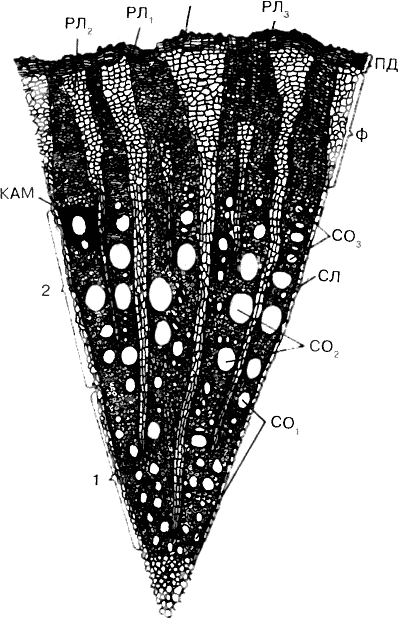
Рис. 6. Анатомическое строение двулетнего корня:
1 — ксилема первого года; 2 — ксилема второго года; Ф - флоэма; ПД — перидерма второго года; кам — камбии; сл — сердцевинные лучи; Рл — радиальные лучи, заложившиеся весной первого года жизни корня; Рл2 — радиальные лучи, заложившиеся весной второго года; Рл3 и Рл4 — радиальные лучи, заложившиеся летом второго года; СО, — сосуды древесины первого года; СО2 и СО3 — сосуды второго года (по Баранову)
В результате жизнедеятельности камбия формируются проводящие пучки, состоящие из отложенных наружу от него сосудов вторичной флоэмы, вовнутрь — вторичной ксилемы. Клетки эндодермы опробковевают, клетки коровой паренхимы и эпидермиса отмирают, что приводит к утере способности к поглощению. Камбий ежегодно весной возобновляет свою работу и функционирует в течение всего вегетационного периода, что приводит к формированию годичных колец. При переходе к вторичному строению из наружных клеток перицикла по всему периметру образуется также кольцо феллогена — пробкового камбия. В процессе деления его клеток наружу откладываются слои пробки, состоящей из мертвых клеток, а вовнутрь — феллодермы, состоящей из живых клеток с целлюлозными оболочками и выполняющей также запасающую функцию.
Пробка, феллоген и феллодерма составляют перидерму — вторичную покровную ткань корня винограда. Ежегодно феллоген закладывается из клеток основной паренхимы в более глубоких слоях флоэмы. В результате формируется корка — защитная ткань из отмерших клеток тканей предыдущих лет перидермы и луба. Из-за ежегодного утолщения корня наружные слои корки разрываются и разрушаются. Пробковый камбий в корнях филлоксеро- и морозоустойчивых форм закладывается глубже, и вторичная пробковая ткань у них более толстая. Представители филлоксероустойчивых североамериканских видов обладают способностью быстро образовывать в местах повреждения филлоксерой раневую меристему и пробковую ткань, изолирующую места повреждения от живой ткани, что не допускает дальнейшего распространения гниения. Такой способностью не обладают представители восточноазиатской и европейско-азиатской групп видов.
Вторичная сосудисто-проводящая система, образующаяся при ежегодном функционировании камбия, состоит из вторичных ксилемы и флоэмы. Вторичная ксилема состоит из проводящих сосудов (трахей и трахеид), древесинной паренхимы и волокон либриформа. По трахеям и трахеидам корня передвигается вода, питательные элементы и другие вещества вверх к стеблю, листьям. Паренхима выполняет запасающую функцию, состоит из живых клеток, расположенных вокруг проводящих сосудов ксилемы. Древесинные волокна, или либриформ, состоят из живых, с заостренными концами, длинных клеток, выполняющих механическую и запасающую функции.
Вторичная флоэма состоит из ситовидных трубок, клеток-спутниц, лубяной паренхимы и лубяных волокон. Ситовидные трубки представляют собой однородные тяжи удлиненных живых клеток, соединенных между собой ситовидными пластинками, имеющими поры. Они вместе с клетками-спутницами обеспечивают передвижение органических веществ. В клетках- спутницах вырабатываются ферменты, которые поступают в ситовидные трубки и регулируют процессы метаболизма. Основная ткань флоэмы — лубяная паренхима (мягкий луб) состоит из тонкостенных клеток, расположенных около ситовидных трубок,
и выполняет запасающую функцию. Лубяные волокна (твердый луб) выполняют механическую и запасающую функции и состоят из плотно прилегающих друг к другу толстостенных клеток склеренхимы. Слои твердого и мягкого луба во флоэме корня винограда чередуются.
Корневая система взрослого куста состоит из многолетних скелетных корней, отходящих от подземного штамба, корней следующих порядков ветвления и молодых корней текущего года. Мощность корневой системы — развитость корней, насыщенность их обрастающими мочками корней, общая площадь поглощающей части — зависит от возраста куста, силы роста куста, ботанического вида, сорта, почвенных условий и др.
Виноградное растение обладает высокой корнеобразующей способностью в сравнении с другими многолетними плодовыми культурами. На процесс корнеобразования влияют видовые и сортовые особенности, состояние черенков, их влажность, запас питательных веществ в них, температура, аэрация и т. д. Важным фактором окоренения является гормональное состояние черенков. Экзогенная обработка черенков регуляторами роста ауксиновой природы способствует стимуляции окоренения. Лучшему окоренению черенков, образованию большего количества корней способствует обработка маточных кустов в период роста побегов (перед цветением) ретардантами. Получению указанного эффекта наряду с изменением гормонального баланса способствует и уменьшение длины междоузлий и сближение узлов побега.
Важнейшим условием успешного выполнения корнем своих функций является его рост. В процессе роста удлиняется корень, перемещается активная поглощающая зона в почвенном пространстве, что создает возможность для выполнения главной функции — всасывания воды и питательных веществ из почвы. В процессе радиального роста (утолщения) формируются элементы проводящей системы, образуется основная паренхима, выполняющая механическую, запасающую и защитную функции. Осевой рост корней винограда осуществляется при развитии верхушечной меристемы — апикальный рост. Рост в толщину происходит в результате жизнедеятельности латеральных меристем — камбия, феллогена, а ветвление — перицикла. Чем сильнее рост и ветвление корня, тем выше интенсивность поглощения.
Так же как и надземная часть, корневая система в течение года имеет два периода: покоя и активной жизнедеятельности, роста. Однако, в отличие от первой, для корней характерно отсутствие физиологического покоя: при наличии благоприятных условий внешней среды они могут расти беспрерывно круглый год.
Рост корней начинается весной, когда температура почвы достигает 6...8о С. Возобновляется активность камбия, апикальной меристемы и перицикла (образуются боковые ответвления). Массовый рост и ветвление корней отмечаются в начале периода формирования листовой поверхности. С повышением температуры до 36о С при достаточной влагообеспеченности рост корней усиливается, при более высоких температурах рост угнетается и корни повреждаются. Оптимальная температура для роста корней — 28...32о С.
В течение вегетации отмечены две волны роста: весенняя и осенняя, причем первая более продолжительная. Ослабление роста корней летом не имеет физиологической, внутренней основы, а связано с недостатком влаги и перегревом почвы. Создание влажностно-термического режима, близкого к естественному (беседочные системы ведения, не допускающие перегрева и пересыхания почвы, достаточная влагообеспеченность), предотвращает летнее падение интенсивного роста.
В процессе новообразования и роста корней потребляется большое количество энергии, которая высвобождается при дыхании. Поэтому для активизации роста корней большое значение имеет достаточная аэрация — доступ кислорода — почвы, рыхлое ее состояние, отсутствие избыточного увлажнения.
Осенью после замедления и прекращения потребления продуктов ассимиляции надземными органами при одновременном сохранении благоприятного водно-термического режима в почве создаются хорошие условия для роста корней. Этот период важен для благополучной перезимовки, создания запасов влаги и питательных веществ в растении, обеспечивающих быстрый рост корней весной. По мере снижения температуры рост корней замедляется и прекращается при 7...8о С.
Поглощение воды и питательных веществ из почвы напрямую связано с интенсивностью роста корней. Перемещение в почвенной среде активной поглощающей зоны в процессе роста корней создает возможности для беспрерывного потребления растениями воды и элементов питания. Следовательно, все, что влияет на рост корней, воздействует на интенсивность поглощения.
Вода поглощается корнями в основном благодаря осмотическому давлению, которое возникает при разнице концентраций клеточного сока эпидермальных клеток корня и почвенного раствора. Поглощенная вода передвигается к центральному цилиндру также благодаря разнице концентраций клеточного сока рядом расположенных клеток. Кроме того, существует и другой путь проведения воды к центральному цилиндру — набухание целлюлозы стенок клеток. Питательные элементы из почвы в эпидермальные клетки поглощающей зоны проникают путем диффузии.
Минеральные соли передвигаются в клетку, где концентрация их ниже, чем в почвенном растворе. Часть неорганических веществ и вода поглощаются и передвигаются активным путем, с затратой энергии, преодолевая давление почвенного раствора. Это еще раз подчеркивает роль дыхания и энергии, выделяющейся при этом, для питания растений.
В передвижении воды и питательных элементов по сосудам ксилемы большую роль играет сосущая сила листьев, которая создается при повышении осмотического давления клеток листьев в результате транспирации и образования в них органических веществ при фотосинтезе. Таким образом, с повышением интенсивности фотосинтеза и транспирации увеличивается скорость поглощения и передвижения воды и питательных веществ. С другой стороны, интенсивное поглощение питательных веществ и воды создает лучшие возможности для синтеза органических веществ. Процессы поглощения и перемещения воды и питательных веществ у винограда происходят интенсивно, что обусловлено наличием высокого корневого давления («плач»), высоким уровнем интенсивности фотосинтеза, транспирации, большим объемом листовой поверхности, хорошо развитой проводящей системой, интенсивностью роста корней и т. д. Отсюда — необходимость создания благоприятных условий воднопитательного режима в слое почвы, где располагается основная масса корней, необходимость регулярного удобрения основными элементами питания.
Архитектоника корневой системы зависит от ряда факторов: возраста, сорта, вида, внешних условии, агротехники. Как показали исследования Н. А. Алиева (1980), в первые годы жизни масса корней намного превышает массу надземной части. С возрастом масса надземной части куста нарастает быстрее и к 4-летнему возрасту становится примерно равной массе корней. Важнейшей задачей ухода за кустами винограда является сохранение баланса массы надземной части и корневой системы.
Угол отхождения придаточных корней зависит от сорта-подвоя. Более вертикальное расположение корней характерно для представителей видов Рупестрис и Берландиери, более горизонтальное — для гибридов Винифера х Берландиери, Берландиери х Рипариа и представителей Рипариа. Гибриды между Винифера х Рупестрис, Рипариа х Рупестрис занимают промежуточное положение.
В одних и тех же условиях в Венгрии сорт Карабурну формировал более короткие и более разветвленные корни, а сорта Италия, Шасла — более длинные и менее разветвленные.
На габитус корневой системы влияют внешние условия: физические свойства и механический состав почвы, близость грунтовых вод и т. д. На песчаных почвах корни проникают глубже. В более северных районах корневая система винограда располагается ближе к поверхности почвы. Избыточная влажность почвы препятствует развитию корневой системы. При достижении грунтовых вод корни поворачивают обратно — наблюдается отрицательный гидротропизм. При наличии плотного непроницаемого слоя в почве корневая система располагается неглубоко.
На глубину залегания и характер развития корневой системы влияют: подготовка почвы, глубина посадки, посадочный материал, особенности ухода за кустами и почвой. Образованию и развитию более мощной корневой системы способствует предпосадочная плантажная вспашка, глубокое рыхление почвы и поддержание ее в этом состоянии, а также катаровка, внесение минеральных удобрений, особенно азота. Кроме того, — создание более крупных форм с большим объемом многолетней древесины. Сильная обрезка приводит к гибели части корней.
Для рациональной подготовки почвы под виноградник и правильного ухода за почвой важное значение имеет изучение архитектоники корневой системы винограда в конкретных почвенных, топографических и других условиях.