Лист — это боковой вегетативный орган виноградного растения, выполняющий ряд важнейших функций. В листе происходит процесс фотосинтеза, т. е. преобразование неорганических веществ в органические. В результате транспирации, осуществляемой листьями, исключается перегрев органов, развивается сосущая сила, обеспечивающая непрерывное поглощение и передвижение воды и питательных веществ в растении. Благодаря дыханию в листьях высвобождается энергия, необходимая для ряда других процессов метаболизма. Через листья осуществляется интенсивный газообмен между растением и окружающей средой.
По происхождению различают листья двух типов: семядольные листья и листья, образующиеся на конусе нарастания побега (настоящие). Семядольные листья образуются только у сеянца винограда. Они закладываются на материнском растении в процессе развития зародыша семени. Семядоли содержат запасы питательных веществ, используемых зародышем при прорастании. При прорастании семядоли выносятся на поверхность субстрата, растут, зеленеют и выполняют все основные функции листа. Семядольные листья сильно различаются по форме, размерам и внутреннему строению от обычных листьев. Все последующие листья сеянца, как и все другие листья винограда, закладываются в результате деятельности конуса нарастания.
Первые 3...14 листьев саженца также закладываются в процессе органогенеза почки на материнском растении, из которой развивается надземная часть. Специфика в развитии этих, так же как и нижних 9... 14 листьев, плодоносных и бесплодных побегов заключается в том, что они перезимовывают в виде зачатка в зимующем глазке. Остальные листья основных побегов и пасынков закладываются и развиваются в текущем году.
Листовой бугорок, отчленившийся на конусе нарастания побега, вытягивается, дифференцируется и формирует листовую пластинку, черешок и два пленчатых сидячих бледно-зеленой окраски прилистника. Прилистники появляются на верхушке побега, располагаются с двух сторон в месте прикрепления черешка к побегу и прикрывают верхушку, выполняя защитную функцию. По мере удаления узла, их несущего, от верхушки прилистники постепенно отсыхают и отпадают.
Лист винограда простой, состоит из черешка и листовой пластинки (рис. 20). Черешком называется суженная стеблевидная часть листа, он выполняет механическую функцию поддержания листовой пластинки и оптимальной ориентации ее по отношению к свету. По проводящей системе черешка осуществляется отток
ассимилятов из листовой пластинки и снабжение ее водой и питательными веществами. В тканях черешка кратковременно сохраняются питательные вещества. Черешок у листа винограда длинный, имеет зеленую или, осенью, различной интенсивности красноватую окраску, опушение. Нижняя часть черешка расширена и в месте прикрепления обхватывает побег с трех сторон. По мере удаления от побега черешок приобретает цилиндрическую форму. В месте прикрепления к листу черешок расширяется и образует со своей верхней стороны углубление в виде желобка. Входя в листовую пластинку, черешок разветвляется и формирует пять основных жилок листа, которые, в свою очередь разветвляясь, образуют густую сеть жилкования. Длина черешка зависит от сорта и условий выращивания. У затененных листьев более длинные черешки.

Рис. 20. Строение листа винограда:
А- средняя (главная) жилка; В и В1 — верхняя пара главных жилок; С и С1 — нижняя пара главных жилок; Б — средняя (центральная) лопасть; Д — нижняя боковая лопасть; Е — верхняя боковая вырезка; Ж — нижняя боковая вырезка; 3- черешковая выемка; а, а1, β, β1 — углы жилкования; К — зубцы на концах лопастей (оконечные зубчики); Л — краевые зубчики; М — дополнительные зубчики; Н — черешок
Листовая пластинка у винограда крупная, расширенная, плоская, по краям зубчатая (рис. 21), разнообразной формы, редко цельная, чаще лопастная. Форма листовой пластинки в зависимости от сорта бывает округлой, яйцевидной, сердцевидной, клиновидной, почковидной.
По рассеченности: цельные, слабо-, средне-, глубокорассеченные (рис. 22), трех- или пятилопастные
(очень редко семилопастные). Лопасти листа отделяются друг от друга боковыми вырезками и черешковой выемкой. Боковые вырезки (рис. 23) и черешковая выемка (рис. 24) у различных сортов различаются по глубине, открытости, виду просвета, характеру дна.
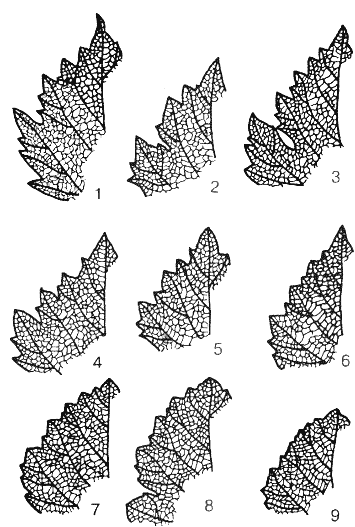
Рис. 21. Типы зубчиков листа винограда:
1 — оконечные зубцы узкотреугольные с оттянутой в острие вершиной, краевые — узкотреугольные и треугольно-пиловидные; 2 — треугольные с широкими основаниями; 3 — оконечные зубцы треугольные с закругленной вершиной, краевые — пиловидные, односторонние выпуклые; 4 — зубчики со слабовыпуклыми сторонами; 5 — зубчики с явно выпуклыми сторонами, но острыми вершинами; 6 — зубчики переходного типа к куполовидным; 7 — куполовидные; 8 — куполовидные с очень широкими основаниями; 9 — очень низкие куполовидные зубчики, зубчатость почти не проявляется
Листовая пластинка состоит из мякоти и жилок. Жилки выступают в виде ребер с нижней, а иногда и с верхней стороны листа. Они выполняют механическую и транспортную функции. В центре располагается главная жилка, которая делит листовую пластинку на две половины. Боковые жилки отходят от места прикрепления черешка к листовой пластинке под определенным углом по отношению друг к другу, называемым углом жилкования. От основных жилок отходят жилки следующего порядка и т. д. Жилки последнего порядка заканчиваются в мякоти листа или входят в зубчики по краям листа. Тип зубчиков по краям листа и на вершинах лопастей используется для ампелографического описания сортов.
Поверхность листовой пластинки винограда бывает гладкой, сетчато-морщинистой, пузырчатой. Она бывает ровной или с изгибающимися вверх или вниз краями. Молодые листья, расположенные на верхушке побега, опушены сильнее, чем взрослые листья, и имеют окраску различных оттенков. Пластинки взрослых листьев винограда могут быть голыми или опушенными, как правило, с нижней стороны. Опушение листьев бывает щетинистым, паутинистым или смешанным (рис. 25).
Окраска листьев изменяется от светло-зеленой до темно-зеленой в зависимости от сорта и условий произрастания. Осенью перед листопадом листья приобретают характерную осеннюю окраску: листья сортов с белыми и розовыми ягодами становятся желтыми, а с черными — красными.
Морфологические признаки листьев изменяются в зависимости от сорта, возраста листа, узла, на котором они располагаются. Наиболее типичные для сорта листья располагаются на 8... 12-м узле.

Рис. 22. Глубина боковых вырезок листа винограда:
1 — мелкая; 2 — средняя; 3 — глубокая

Рис. 23. Типы боковых вырезок листьев винограда:
А — закрытые боковые вырезки:
1 — почти без просвета 2 — с узкоэллиптическим просветом; 3 — с широким эллиптическим (овальным) просветом; 4 — с яйцевидным просветом и заостренным дном; 5 — с яйцевидным просветом и округлым дном; 6 — с треугольным просветом и плоским дном; 7 — с треугольным просветом и однозубчатым дном; 8 с поперечно-овальным просветом
Б — открытые боковые вырезки:
1 — едва намеченные; 2 — в виде входящего угла; 3 — щелевидные; 4 — лировидные с узким устьем и острым дном; 5 — лировидные с узким устьем и округлым дном; 6 — лировидные с узким устьем и плоским дном; 7 — с параллельными сторонами и острым дном; 8 — с параллельными сторонами и округлым дном; 9 — с параллельными сторонами и плоским дном; 10 — с однозубчатым дном
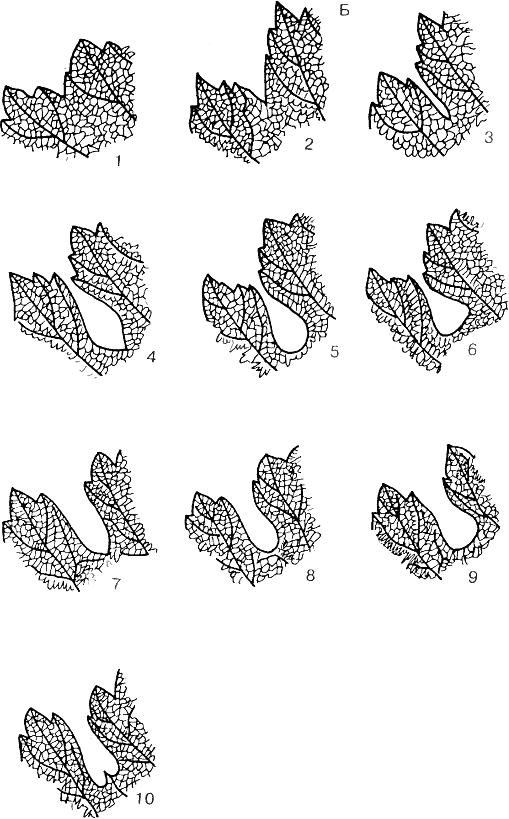
Рис. 24. Типы черешковых выемок листьев винограда:
А — закрытые: 1 — почти наглухо закрытые; 2 — с просветом щелевидной формы; 3 — с просветом эллиптической формы; 4 — с просветом округлой формы; 5 — с просветом яйцевидной формы и округлым дном; 6 — с просветом яйцевидной формы и острым дном; 7 — с просветом обратнояйцевидной формы; 8 — с просветом поперечно эллиптической формы; 9 — с просветом поперечно-эллиптической формы и дном, ограниченным жилками.
Б — открытые лировидные: 1 — с округлым дном; 2 — с острым дном; 3 — с плоскозаостренным дном; 4 — с дном, ограниченным жилками


В — открытые сводчатые:
1 — узкие с округлым дном; 2 — узкие с острым дном; 3 — широкие с плоскозаостренным дном.
Г — открытые стрельчатые:
1 — узкие; 2 — равносторонние; 3 — очень широкие; 4 — с двумя сложными шпорцами
Лист винограда относится к органам с ограниченным ростом, т. е. растет в течение 5...6 недель, достигая определенного размера, после чего рост его прекращается. По мере роста усиливается процесс ассимиляции, изменяется химический состав листа. Осенью активность ассимиляции падает, хлорофилл разрушается, происходит отток питательных веществ из листа в стебель, листья приобретают осеннюю окраску. В месте прикрепления черешка листа к побегу закладывается пробковый отделительный слой и листья опадают. На месте соединения черешка побега остается листовой след. Если поверхность листового следа гладкая и имеет коричневую окраску, это значит, что листья закончили вегетацию, побеги вызрели нормально. Шероховатая поверхность и серо-грязная или черная окраска листового следа свидетельствуют о том, что листья не завершили вегетацию вследствие затягивания ее или заморозков и побеги плохо вызрели.
Анатомическое строение черешка, жилок и листовой пластинки соответствует выполняемым ими функциям. В черешке хорошо развиты проводящая система и механическая ткань. Проводящая система черешка состоит из расположенных в виде колец проводящих пучков, между которыми находятся сердцевинные лучи. Большое количество пучков, сосудов и ситовидных трубок создает возможности для интенсивного снабжения листовой пластинки водой, элементами питания и быстрого оттока продуктов ассимиляции. Успешному выполнению механических функций способствует наличие кольца первичной механической ткани — колленхимы — и перицикловых склеренхимных волокон. В черешке хорошо развиты коровая и древесинная паренхимы, создающие возможность для временного хранения запасных питательных веществ при их передаче в лист или в стебель. Снаружи черешок, так же как и жилки, покрыт эпидермисом и кутикулой. Главные жилки, которые образуются черешком при входе в листовую пластинку, имеют подобное же строение, только меньшее количество проводящих пучков. В жилках последующих порядков, при сохранении общей анатомической структуры, последовательно уменьшается размер и количество проводящих пучков.
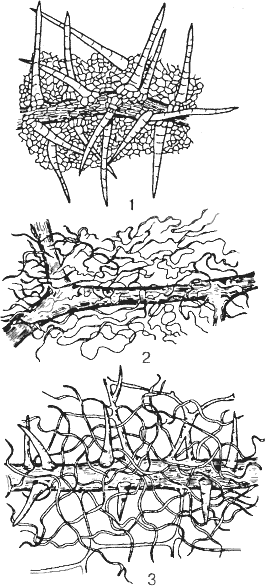
Рис. 25. Типы опушения листьев винограда:
1 — щетинистое; 2 паутинистое; 3 — паутинисто-щетинистое
В межжилковом пространстве анатомическое строение листа имеет обычную структуру (рис. 26). С верхней и нижней стороны лист покрыт эпидермисом, состоящим из одного слоя плотно прилегающих друг к другу клеток, снаружи которых образуется кутикула. Эпидермальные клетки черешка и листовой пластинки, особенно с нижней ее стороны, образуют выросты (волоски), составляющие опушение различных типов. На нижней стороне листовой пластинки (на верхней очень редко) клетки эпидермиса образуют устьица, обеспечивающие регулируемый обмен с окружающей средой. Их количество на единицу площади листа в зависимости от сорта, возраста листа, места на побеге, условий формирования (влажность, освещенность и др.) колеблется от 120 до 200 шт. на 1 мм2 . На концах зубчиков листьев формируются водные устьица (гидатоды), которые способны выделять воду в жидком виде. Этот процесс наблюдается при низкой транспирации, высокой влажности (часто встречается в теплицах), при хорошей влагообеспеченности и на молодых листьях и называется гуттацией.
Между верхним и нижним эпидермисом располагается мякоть листа — мезофилл, имеющий дорзивентральное строение. К верхнему эпидермису прилегает слой клеток палисадной, или столбчатой, паренхимы, вытянутых перпендикулярно поверхности листа, богатых хлорофиллом в хлоропластах и обеспечивающих, в основном, фотосинтез. Под ним располагается слой собирательных клеток, вытянутых в том же направлении, но более коротких и обеспечивающих передачу продуктов ассимиляции проводящей системе жилок, которые своими окончаниями прилегают к ним.
Под этим слоем, примыкая к нижнему эпидермису, располагается губчатая ткань, состоящая из 4...7 слоев неправильной округлой формы клеток с крупными межклетниками. Клетки губчатой ткани содержат небольшое количество хлоропластов и выполняют функцию газообмена и также передают ассимиляты в проводящую систему жилок.
Листья винограда характеризуются очень высоким уровнем важнейших процессов жизнеобеспечения растительного организма — фотосинтеза, транспирации, дыхания. В результате деятельности отдельных листьев формируется совокупная биологическая и хозяйственная продуктивность побега, виноградного куста и насаждения. Отдельные листья при этом функционируют в условиях не характерного для естественно произрастающего растения, а создаваемого искусственно ценоза.

Рис. 26. Анатомическое строение листа:
1 — кутикула; 2 — эпидермис верхней стороны листа; 3 — эпидермис нижней стороны листа; 4 — палисадная ткань; 5 — слой собирательных клеток; 6 — губчатая ткань; 7 — устьице; 8 — хлорофилловые зерна; 9 — проводящий пучок; 10 — сферокристалл; 11 — рафида; 12 — межклетники
Листовая поверхность
Листовой поверхностью называется суммарная площадь листьев побега, куста, ряда и виноградника. Величина листовой поверхности, ее структура, условия ее функционирования определяют величину биологического и хозяйственного урожая, качество продукции. Существуют различные методы определения величины листовой поверхности: весовой, планиметрический, расчетный (ампелометрический). Сущность наиболее распространенного в виноградарстве ампелометрического метода заключается в том, что площадь листа рассматривается как площадь круга с диаметром, равным условному диаметру листа (от верхнего зубца на центральной лопасти до наиболее удаленного зубца нижней боковой лопасти). Затем вносится поправочный коэффициент на рассеченность листовой пластинки, который составляет для цельных листьев 1,25; трехлопастных — 1,27; для пятилопастных слаборассеченных — 1,30 и си льнорассеченных — 1,35.
Листовая поверхность побега достигает 16... 18 дм2 и больше, куста — 11... 14 м2 , гектара виноградника — 35...40 тыс. м2 . Наиболее существенно на этот показатель влияет нагрузка побегами, форма, уход, влагообеспеченность и др.
Однако интенсивность фотосинтетической деятельности, интенсивность и направленность других важных процессов метаболизма определяются архитектурой куста — расположением листовой поверхности в пространстве и развитием ее во времени (фотосинтетический потенциал). При оценке кроны куста с точки зрения поглощения ФАР ее моделируют как призму, цилиндр и другие геометрические фигуры. Естественно, что листья, расположенные на разных ярусах, различных слоях, получают разную величину ФАР и характеризуются разной продуктивностью. Индекс листовой поверхности рассчитывается как отношение односторонней площади листьев к площади поверхности земли, которую эта листовая масса занимает. Показатель является весьма важным для характеристики величины листовой поверхности, условий функционирования листьев. Для этих же целей используют показатель плотности площади листьев — отношение площади листьев к объему кроны, в которой они располагаются (м2 /м3 ). Необходимо иметь в виду, что оптимальные величины этих показателей для крон различной архитектуры (штамбовые, бесштамбовые, способ ведения прироста, плотность посадки) неодинаковы.
Важнейшим показателем, характеризующим листовую поверхность винограда, является фотосинтетический потенциал — сумма. ежедневных площадей листьев куста, ряда или насаждения за период формирования урожая или часть этого периода. Разным величинам фотосинтетического потенциала каждого сорта соответствует разная величина биологического и хозяйственного урожая. Так, для получения 100 ц/га урожая гроздей сорта Ркацители необходимая расчетная величина фотосинтетического потенциала составляет 1,26...1,76·106м2 ·день, а для получения 150 ц/га — 1,83...2,64·106м2 ·день.
Листовая поверхность куста и насаждения нарастает постепенно: максимальная величина ее достигается к началу созревания, к началу цветения — 15...30% от нее, а через 20...25 дней после цветения — 60...65% от максимальной.
Фотосинтез
Интенсивность фотосинтеза оценивается через количество углекислоты, ассимилированной листьями. Этот показатель фотосинтетической деятельности виноградного растения зависит от ряда внешних и внутренних факторов: освещенности, температуры, влагообеспеченности, поступления питательных веществ и углекислоты, сорта, вида, возраста листьев, возраста кустов, агротехнических приемов и др.
Фотосинтез возможен только на свету. Компенсационная точка светового фактора, т. е. величина освещенности, когда количество углекислоты, поглощаемой при фотосинтезе и выделяемой при дыхании, одинаково, в зависимости от сорта и вида колеблется в пределах 300... 1 500 лк. Наиболее низкий уровень компенсационной точки характерен для вида V. riparia. По мере увеличения освещенности увеличивается и интенсивность фотосинтеза — эта зависимость имеет вид гиперболической кривой. Световая насыщенность фотосинтеза у винограда разных видов наступает при разных уровнях освещенности, начиная с 30...40 тыс. лк. Несмотря на невысокие показатели светового насыщения, необходимо иметь в виду, что по мере удаления листьев от поверхности в глубь кроны значительно снижается освещенность. У затененных листьев внутри кроны при загущении не только слабая интенсивность фотосинтеза, но они тратят большее количество пластических веществ на дыхание, особенно при повышенных температурах. Поэтому важны приемы, регулирующие световой режим виноградника.
Оптимальной температурой для фотосинтеза является 28...32° С. При температуре выше 35° С и ниже 6° С фотосинтез прекращается.
Важнейшим фактором фотосинтеза является и водный режим. При недостатке влаги интенсивность ассимиляции резко падает. В засушливые годы в растении плохо накапливаются запасные и защитные вещества, медленно созревают ягоды, плохо вызревают побеги, в результате снижается морозо- и зимостойкость растений. При относительной влажности воздуха ниже 50% и влажности почвы ниже 45% Ш1В существенно снижается интенсивность фотосинтеза.
На интенсивность фотосинтеза влияет концентрация углекислоты в воздухе. С повышением концентрации СО2 до 0,1% происходит крутой подъем интенсивности фотосинтеза, дальнейшее ее повышение, однако, менее эффективно. Отмечено также, что повышение концентрации СО2 резко усиливает эффективность использования влаги. Внесение в почву виноградника органических удобрений способствует увеличению концентрации СО2 в приземном слое воздуха и стимулирует фотосинтез.
На интенсивность фотосинтеза влияют и элементы минерального питания. Недостаток или избыток некоторых из них может тормозить фотосинтез.
Для сохранения интенсивности фотосинтеза на высоком уровне необходим постоянный отток продуктов ассимиляции из листьев. Поэтому на кустах с увеличением нагрузки урожаем интенсивность фотосинтеза повышается. И, наоборот, с увеличением листовой поверхности интенсивность фотосинтеза снижается. Поэтому листья плодоносных побегов отличаются более высоким уровнем фотосинтеза.
В фотосинтезе листьев винограда существует суточная и сезонная динамика. В более теплых районах виноградарства в течение дня наблюдаются два пика интенсивности фотосинтеза: первый пик — в дополуденные часы, затем — спад в середине дня и второй пик — после полудня. Установлено, что для листьев разных ярусов пики интенсивности наблюдаются в разное время дня.
Фотосинтетическая активность листьев разных ярусов на побеге меняется в течение вегетации, зона максимальной интенсивности фотосинтеза смещается в акропетальном порядке. Меняется и направление перемещения ассимилятов от листьев разных ярусов. Количество листьев, работающих для нисходящего оттока ассимилятов, увеличивается: перед цветением один лист, расположенный выше соцветия, направляет ассимиляты вниз, после цветения — четыре, в начале созревания ягод — девять и т. д. К периоду созревания нижние 3...4 листа практически прекращают снабжать грозди углеводами, вместе с тем их наличие ухудшает микроклимат в зоне гроздей. Поэтому рекомендуется их в этот период удалять.
Благодаря фотосинтезу нижних 8...10 листьев формируется масса гроздей, а на вышерасположенных лежит накопление сахаров и формирование качества. Из этого можно сделать вывод: для накопления сахаров имеют значения листья пасынков. Все изложенное необходимо учитывать при проведении операций с зелеными частями куста.
Дыхание
Дыхание — важный процесс метаболизма, в ходе которого сложные органические соединения окисляются, в результате чего выделяется свободная энергия, используемая для роста, развития и т. д. В результате дыхания образуются углекислый газ и вода. Интенсивность дыхания зависит от ряда внешних и внутренних факторов. С повышением температуры интенсивность дыхания листьев возрастает, максимальная ее величина наблюдается при 40° С. При температуре, повреждающей протоплазму, дыхание снижается. Интенсивность дыхания молодых листьев в несколько раз выше, чем взрослых. Установлены также изменения в интенсивности дыхания листьев в течение вегетации: в начале вегетации она нарастает, достигает максимума в период массового цветения, затем снижается к листопаду. Средняя величина интенсивности дыхания для листьев винограда составляет 1,5...2,5 мг/дм2 ч.
Одной из важнейших функций листа является транспирация — процесс испарения воды. Вода испаряется в основном через устьица (устьичная транспирация), а в небольшом объеме — через кутикулу (кутикулярная транспирация). Потеря воды клетками листа приводит к возникновению сосущей силы, которая, передаваясь от клетки к клетке, затем — к сосудам древесины и далее к корням, обеспечивает непрерывное поглощение и подачу воды и питательных веществ в листья.
Другая важная роль транспирации — предотвращение перегрева листьев, а также других органов. Поступающая в листья вода также участвует в процессе фотосинтеза. Вместе с тем чрезмерно интенсивная транспирация при недостатке воды в почве приводит к завяданию листьев.
Виноградному растению присуща высокая интенсивность транспирации. Так, в опытах с сортом Рислинг установлено, что листья площадью в 1 м2 транспирируют 1,9...2,0 г воды в минуту. Один куст, имеющий 150...200 листьев, при температуре 24о С испаряет 1,0...1,5 л, а виноградник площадью 1 га с общей листовой поверхностью 30 000 м2 — 30 т воды в сутки. Транспирационный коэффициент винограда составляет 120...400 л на 1 кг сухого вещества. Из общего количества поглощаемой растением винограда воды на формирование биомассы уходит 0,2%, остальное расходуется на транспирацию.
Транспирация имеет определенную суточную динамику: в утренние часы интенсивность ее повышается и достигает максимума в около полуденные часы. Одними из важнейших факторов, определяющих величину транспирации, являются температура и влажность воздуха, а также влажность почвы. Резко повышает транспирацию сухой воздух, ветер. На 2...7-й день после полива у орошаемых кустов транспирация в 1,5...2,5 раза выше, чем у неорошаемых кустов. Установлены видовые и сортовые различия в интенсивности фотосинтеза: V. vinifera обладает самой высокой транспирационной активностью, а V. riparia — самой низкой.
Виноградное растение способно регулировать интенсивность транспирации благодаря способности устьиц активно закрываться. При недостатке влаги устьица закрываются, транспирация ограничивается. Одновременно снижается интенсивность ассимиляции, так как уменьшается поступление углекислоты из атмосферы.