При наличии благоприятных условий внешней среды возбудитель милдью стремится осуществить бесполое размножение. Оно заключается в том, что сплетение грибницы паразита, заполняющее собой подустьичные полости в пределах зараженной ткани, начинает быстро развивать конидиеносцы, выходящие через отверстия устьиц на поверхность зараженных листьев. Скопление таких конидиеносцев образует характерный для милдью снежно-белый пушок, выступающий на нижнюю поверхность листьев. Конидиеносцы при своем формировании отделяются от мицелия клеточной перегородкой.
Длина конидиеносцев может сильно варьировать в зависимости от влажности и температуры окружающего воздуха, а при благоприятных условиях конидиеносцы достигают 1 мм длины. Через каждое устьице на поверхности пятна милдью в среднем выходят 3—6 конидиеносцев. Однако в отдельных случаях их количество может достигать 20 на одно устьице. Конидиеносцы моноподиально ветвятся, обычно образуя от 2 до 7 боковых ветвей, на каждой из которых возникают вторичные ответвления (2—4 шт.), которые в свою очередь ветвятся, образуя короткие стеригмы, на которых развиваются летние зооспорангии (рис. 15). Следут отметить, что интенсивность ветвления конидиеносцев, количество и размеры развивающихся на них зооспорангиев весьма варьируют в зависимости от погодных условий. При благоприятных условиях внешней среды для бесполого размножения паразита летние зооспорангии — «микроконидии»—приобретают овальную форму, бесцветны и имеют размеры 14—25ц. X 10—16р, в среднем 20 X 14р. Помимо факторов внешней среды, влияющих на величину зооспорангиев и конидиеносцев, аналогичное влияние могут оказывать видовые и сортовые различия винограда. У наиболее восприимчивых видов и сортов при благоприятных условиях среды формируются конидиеносцы обильно ветвящиеся, а у более устойчивых—со слабым ветвлением. Т. Сэвулеску (1941), проводя сравнительные измерения величины зооспорангиев возбудителя милдью у восприимчивого европейского винограда V. vinifera и у устойчивого дикого декоративного винограда Ampelopsis quinquefolia, получил следующие различия в размерах зооспорангиев:
Vitis vinifera 18,9ц,X 15,15р Ampelopsis (Parthenocissus) qninguefolia 20,33ц,X 15,93р. Соответствующим образом изменена и форма конидиеносцев (рис. 16). Следует отметить, что образование конидиеносцев с зооспорангиями наблюдается в природе только в ночные часы, когда у растений прекращается фотосинтез. Искусственно вызвать образование конидиеносцев днем можно, только погрузив или целое растение или отдельные сорванные зараженные листья в темноту в условия высокой влажности на продолжительное время (3—5 час). Работами Раваза (1911) и К Мюллера (1923) установлено, что минимальной температурой, при которой происходит массовое образование конидиеносцев, является 12—13°. При наличии минимальных ночных температур выше 13° создаются температурные условия, благоприятные для спороношения возбудителя милдью.

15. Образование летних зооспорангиев на конидиеносцах возбудителя милдью. Формирование и выход зооспор из зооспорангиев (по Т. Сэвулеску).
Образование конидиального спороношения (зооспорангиев) возможно, однако, и при более низких температурах. Так, Иштванфи и Палинкас (1913) наблюдали его возникновение даже при 8°. Позднее А. Цеснер (A. Zeesnert) также установил возможность появления спороношения PI. viticola при 9,5°. А. Д. Липецкая (1931) наблюдала также возникновение на поверхности листьев конидиального спороношения милдью даже в условиях температуры 7,6°. Мы также неоднократно наблюдали возникновение у PI. viticola бесполого спороношения при температурах около 10°. Однако массовое образование зооспорангиев в природе происходит при температурах выше 12°—13°.

16. Изменение формы конидиеносцев под влиянием питающего растения. Справа — у Vitis vinifera, слева — у Ampelopsis quinquefolia (по Т. Сэвулеску).
Оптимальными температурами являются 20—25°, а максимальной — 29°. Условия влажности, необходимые для обильного спороношения согласно данным Т. Сэвулеску, должны превышать 95% относительной влажности воздуха, хотя слабое спороношение возможно уже при относительной влажности воздуха 92%. Практически в условиях природы такого рода высокая влажность в ночные часы сопровождается выпадением росы или образованием тумана. Поэтому в природе спороношение паразита происходит при наличии увлажнения листвы винограда дождем, росой или туманом.
Образующиеся на конидиеносцах зооспорангии очень легко отделяются от стеригм благодаря образованию отделяющего слоя (перегородки), состоящего из коллозы (P. Viala, 1901). При малейшем движении воздуха, при наличии влажности перегородка из коллозы набухает, ослизняется, а зооспорангии легко сдуваются ветром и могут переноситься на значительные расстояния.

17. Влияние температуры на быстроту прорастания зооспорангиев возбудителя милдью.
Оседая из воздуха на влажные листья и другие зеленые органы винограда, зооспорангии растрескиваются, освобождают находящиеся в них зооспоры, и в дальнейшем начинается развитие новой генерации паразита путем такого же заражения растений, как это происходит при первичной инфекции от ооспор.
Быстрота прорастания летних зооспорангиев (выход зооспор из спорангия) в сильной степени зависит от температуры. Выяснению этого вопроса были посвящены исследования ряда авторов: Раваз (1914), Мюллер и Шлеумер (1934), Мейер (1941), Штеллвааг (1954). Результаты исследований приводятся » графиках на рис. 17. Из графиков видно, что все указанные исследователи пришли к выводу, что в диапазоне температур от 15° до 25° прорастание зооспорангиев, погруженных в воду, происходит в течение около 1 часа. С понижением температуры прорастание зооспорангиев резко замедляется. По Штеллваагу, Мюллеру, Шлеумеру и Майеру, при 5° оно достигает 10—12 час, а по Равазу, при температурах ниже 10° приближается к 20 час. При повышении температур выше 25° выход зооспор из зооспорангиев также замедляется и при температурах выше 29° не происходит вообще.
Согласно данным Гоймана (1950) и Штеллваага (1954), при температуре, равной 30°, во влажном воздухе в спорангиях не происходит формирования зооспор, и они могут прорастать непосредственно ростковой гифой, как конидии. Д. Бубалс (1958), пытаясь проверить эти данные, не смог наблюдать подобного рода превращения зооспорангиев в конидии.
По данным Т. Сэвулеску, на прорастание зооспорангиев влияет рН воды: при рН от 4 до 8 прорастание происходит нормально; при рН ниже 4 не происходит выхода из спорангиев зооспор; при рН выше 8 прорастание спорангиев происходит без образования зооспор, путем выхода наружу протоплазматической массы, лишенной жгутиков.
Branas, Bernon и Bellet (1942) проращивали спорангии в дождевой воде с рН, равным 5,2.
Наши опыты по действию рН среды на жизнеспособность зооспор Pf. viticola также подтверждают данные Branas, Bernon и Bellet.
Как видно из таблицы 11, выход зооспор из зооспорангиев и движение наблюдается при рН, равном от 7 до 5,5.
Таблица 11
Действие рН среды на жизнеспособность зооспор Plasmoparaviticola
| рН среды | Выход зооспор из зооспорангиев | Движение зооспор |
РН 7,0 |
| Хороший | Интенсивное |
РН=6.5 | (подкисленная |
|
|
| лимонной кислотой) | Хороший | Интенсивное |
РН =6,о |
| Хороший | Интенсивное |
РН-5,5 |
| Единичный | Слабое |
РН-5,0 |
| Нет | Нет |
РН- 4,5 |
| Нет | Нет |
РН =4,0 | „ | Нет | Нет |
РН -=3,0 |
| Нет | Нет |
Gaumann (1950) сообщает, что дождевая вода на поверхности листьев обычно приобретает слабощелочную реакцию с рН = = 7,2 — 7,8 вследствие диффузии солей во время транспирации растений.
Следует отметить, что при образовании маслянистых пятен милдью на сравнительно молодых листьях клетки больной ткани обычно долго остаются живыми (до 15—20 дней). В связи с этим при наличии благоприятной погоды (теплые и влажные ночи) образование новых конидиеносцев может происходить многократно. Однако интенсивность их повторного образования зависит от степени расходования мицелием гриба, находящимся внутри пораженных листьев, накопленных им ранее запасных питательных веществ. Если первое спороношение происходило при вполне благоприятных условиях внешней среды и было весьма обильным, то повторение спороношения бывают более слабыми, а развивающиеся при этом зооспоры обладают меньшей продолжительностью своего движения. Обычно при наличии благоприятных условии спороношение гриба на зараженных листьях повторяется 7—8 раз. Причем наибольшей вирулентностью обладают зооспоры, сформировавшиеся при первом наступлении вполне благоприятных условий для спороношения.
Таблица 12
Способность к образованию повторных спороношений грибом Plasmopara viticola, 1933 год
| Даты спороношений | Последовательность спороношений | Число повтор-ностей | Среднее количество зооспорангиев |
15/УП | 1-е конидиальное спороношение . | 100 | 51,76± 1,402 |
17/УП | 2-е конидиальное спороношение | 100 | 28,46± 1,022 |
18/УП | 3-е конидиальное спороношение . | 100 | 24,05 ±0,821 |
20/УП | 4-е конидиальное спороношение . | 100 | 19,0 ±0,662 |
22/УП | Б-е конидиальное спороношение . | 100 | 6.67 ±0,248 |
25/УП | 6-е конидиальное спороношение . | 90 | 9,35 ±0,289 |
26/УП | 7-е конидиальное спороношение . | 60 | 3,62 ±0,209 |
28/УП | 8-е конидиальное спороношение . | 100 | 5,39 ±0,166 |
Д. Д. Вердеревский (1953) сообщает о результатах специальных опытов, проведенных им совместно с А. Т. Макрушиной по проверке интенсивности образования повторных спороношений на одних и тех же пятнах милдью, согласно которым при благоприятных температурных условиях для спороношения основная масса зооспорангиев образуется в течение первых двух ночей, хотя слабые повторные спороношения наблюдаются в течение пяти суток. При худших температурных условиях сравнительно обильные новые спороношения наблюдаются в течение четырех ночей и способность к спорообразованию растягивается на шесть суток. Э. Б. Райков (1958) подтвердил эти данные в условиях Болгарии.
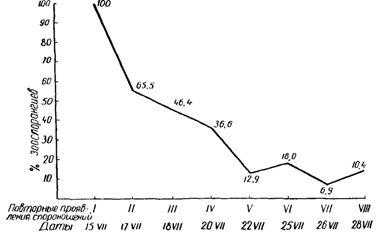
18. Снижение интенсивности спороношения возбудителя милдью при повторных проявлениях болезни на одних и тех же пятнах милдью. (В процентах от первого спороношения, сорт Шасла белая, 1966 г.).
Нами в 1966 году также была проверена способность возбудителя милдью к повторному спороношению при температурах + 20°, +25° в природных условиях на отдельных кустах сорта Шасла. Результаты этого опыта приведены в табл. 12 и в графике на рис. 18.