X. Дюринг
ФИТОГОРМОНЫ ВИНОГРАДНОЙ ЛОЗЫ
ВВЕДЕНИЕ
Понятие фитогормоны охватывает 5 групп фитогенных веществ, передвигающихся в растении и в незначительных дозах индуцирующих или контролирующих различные физиологические процессы. Обозначенные как "химические сигналы", гормоны в большей части обладают высокой подвижностью во всем растении, так что регулирующие функции этих веществ проявляются в месте их синтеза или, в известной степени при коррелятивных отношениях, в других органах. В более узком смысле к фитогормонам причисляются ауксины, гиббереллины, цитокинины, абсциссины и этилен. В разделе, где описываются выявляемые аналитическим путем фитогормоны виноградной лозы, речь идет и о дополнительных веществах, оказывающих ингибирующее действие, которые использовались в биологическом тексте. В следующем разделе рассматриваются вопросы действия синтезируемых аналогов фитогормонов и аналогичных им веществ, напоминающих фитогенные гормоны.
Исследовательская работа в целях определения гормонов виноградной лозы и их действия в последние годы разрослась до такой степени, что в объеме столь небольшой работы не представляется возможности охватить новейшую литературу по этому вопросу примерно после середины 60-х годов. Данные по этому вопросу можно найти в следующих обзорах: Аllеwеldt с сотр. (1967, а, b, 1975, 1977), Coombe (1973, 1976а), During, Allewеldt (1974), During (1976a), Hidalgo, Candella (1973), а также в диссертациях Lott (1969), Hifnу (1971), During (1972), Juliiard (1973), Waitz (1975), Luckert (1976), Vollmer (1976), Koch (1977).
ЛОКАЛИЗАЦИЯ ФИТОГОРМОНОВ
Фитогормоны в вегетативных органах
Ауксины. Сезонная ритмика фаз роста и покоя виноградной лозы дает основание считать, что стимулирующие и подавляющие рост вещества участвуют в регулировании роста в отдельные времена года. Исследования Spiegel (1954) и Мashеvа (1975) показывают, что вещества, проявляющие ауксинную активность в биологическом тексте с побегами и почками, доказуемы, причем наиболее высокое их содержание установлено до распускания почек, после полного исчезновения растворимой в эфире ингибирующей фракции.
Эти результаты подтверждаются газохроматографическими анализами I s о d а (1975). Lilov и Nikolova (1974) установили, что физиологически молодые части побега, как и находящиеся под ними листья, отличаются большим содержанием ауксинов по сравнению с более старыми зонами побегов и листьями под ними. В главных и боковых корнях содержание ауксинов в течение вегетационного периода повышается, а после наступления зимнего покоя понижается. Наблюдавшийся Masheva (1975) базипетальный транспорт ауксинов свидетельствует в пользу предположения, что эти гормоны во время вегетационного периода передвигаются в корнях (Lilоv, Niкоlоvа, 1974).
Цитокинины. Активность цитокининов в виноградной лозе была впервые доказана Nitsch и Nitsch (1965) в пасоке, а впоследствии, при применении того же метода, и в листьях (J а к о, 1970, 1977). V о ll m е г (1976) подробно исследовал годичный ритм цитокининов в вегетативных органах разных сортов Vitis vinifera и на различных подвоях в условиях теплицы и открытого грунта. Низкое содержание их в зимних глазках в фазу эндогенного покоя сопровождается повышением активности в период после покоя до прорастания почек. До наступления сокодвижения это повышение, очевидно, объясняется синтезом или активацией цитокининов в почках, после чего наступает дополнительное накопление цитокининов, вероятно, из корней. Максимальное содержание цитокининов, установленное во время прорастания почек, сопровождается снижением его в молодых побегах до минимальных значений через 4—6 недель после прорастания. В июне содержание цитокининов вновь повышается в более старых листьях, тогда как в побегах и зимних глазках их содержание уменьшается с наступлением осени. Наподобие листьев, и в корнях установлено увеличение цитокинина во время вегетационного периода; более молодые части корней, однако, богаче цитокинином, чем более старые. Конус нарастания побегов также содержит больше цитокинина, чем листья. У всех исследовавшихся сортов и сочетаний прививок установлено повышение концентрации цитокинина по мере усиления интенсивности сокодвижения до прорастания почек и понижение — с ослаблением интенсивности плача после распускания почек. Очевидно, концентрация цитокинина в пасоке определяется преимущественно привоем.
Исследования Lilov и Andonowa(1976) подтверждают эти результаты. Во время роста ягод наступает снижение содержания цитокининов в листьях, а во время их созревания оно вновь повышается. Примерно до июля в побегах и корнях устанавливается повышение цитокинина, после чего его активность начинает снижаться до середины ноября, а затем опять повышается в декабре. Итак, за исключением листьев, во время эндогенной фазы покоя во всех органах виноградной лозы устанавливается незначительная, а впоследствии в различной степени возрастающая активность цитокининов.
Исследования в связи с повышением содержания цитокининов при различной температуре показали, что хранение в условиях низкой температуры (+1°С) повышает активность цитокининов в вызревших побегах (Skene, 1972), а также, что разная температура питательного раствора приводит к изменению содержания отдельных цитокининовых фракций в корневом экссудате (Skene и Keridge, 1967). Нарушение соотношения источник — потребитель (source — sink) у плодоносящих кустов путем удаления гроздей понижает активность фотосинтеза и в то же время усиливает образование цитокининовых глюкозидов в листьях (Hoad с сотр., 1977). Повышение содержания цитокинина примерно в 20 раз установлено в пасоке, когда в питательный раствор добавляли хлорхолинхлорид (ССС) (Skene, 1968, 1970).
Наоборот, снижение активности цитокинина в корнях отмечалось при уменьшении доз N или К. В условиях без удобрения NPK в корнях не обнаруживалось цитокининов (Jако, 1974).
Гиббереллины. До прорастания почек Isоdа (1973) установил повышение активности гиббереллинов (GA) в почках. В корневом экссудате и в экстрактах из листьев и корней Skene (1967) обнаружил вещества, Rf- значения и биологическая активность которых соответствовали GA4 и GA7. В усиках установлено большее содержание гиббереллина, чем в других органах побега; закручивание усиков сопровождается ростом гиббереллинов (Маnanкоv, 1976). По мнению Kocherzhenko и Maiko (1967), между содержанием гиббереллина в листьях и ростом существует тесная корреляция. Так, например, в условиях продолжительного освещения и длинного дня в листьях быстрорастущих растений при помощи теста прорастания гороха установлены гиббереллины, которых не обнаружено в условиях короткого дня при редуцированном росте. Удаление гроздей, по данным Hoad с сотр. (1977), ведет к уменьшению гиббереллиноподобных веществ в листьях.
Абсциссины и ингибиторы. Впервые в пасоке виноградной лозы при помощи биологического теста Rapp и Ziegler (1971)установили абсциссиновую кислоту на основе Rf-значений в различных жидкостях, поведения в ультрафиолетовой области, а также инфракрасного и массового спектров. На основе этих методов впоследствии были исследованы сезонные изменения абсциссиновой кислоты в побегах и листьях: содержание абсциссиновой кислоты в листьях, где она синтезируется, не претерпевает существенных изменений в течение вегетационного периода. В побегах, наоборот, обнаруживается четкая сезонная динамика, причем максимум устанавливается в эндогенную фазу покоя (During, Alleweldt, 1973; Cheng с сотр., 1974). Решающим фактором повышения содержания абсциссиновой кислоты можно считать уменьшение длины дня, что, однако, не снижало ее количества в листьях, подвергнутых раздражению фотопериодом. Взамен этого, уменьшение длины дня увеличивает абсциссиновую кислоту в побегах, что свидетельствует о ее передвижении из листьев к побегам (Alleweldt, During, 1972). В апикальных частях побегов нередко устанавливалось большее содержание абсциссиновой кислоты, чем в секторах у основания (Lilov, Angelova, 1977а; During, неопубликованные данные). Как показывают опыты Lоveys и Kriedemann (1973), наблюдается быстрая и четкая реакция содержания абсциссиновой кислоты в листьях на продолжительный водный дефицит; при пониженной интенсивности транспирации и фотосинтеза, а также при повышении водного потенциала, содержание абсциссиновой кислоты в листьях увеличивается, тогда как после повторного полива наблюдается обратное. Наряду с повышением содержания абсциссиновой кислоты в листьях увядающих виноградных кустов устанавливается еще большее его повышение в побегах (During, Sсiеnz а, 1975), что, по всей вероятности, приводит к прекращению роста побегов в длину. Исследования показали, что суточный ход транспирации в большинстве случаев находится в отрицательной корреляции с содержанием абсциссиновой кислоты в листьях (During, Scienza, 1975). Из листьев выделена также близкая абсциссиновой фазеиновая кислота (Loveys, Kriedemann, 1974).
В нейтральных, основных и кислых экстрактах, из побегов Тiziо с сотр. (1968) установили при помощи биологического теста ингибиторы. Weaver с сотр. (1968) и Sarkisova с сотр. (1976) установили в начале периода покоя повышение феноловых ингибиторов в побегах и почках, содержание которых уменьшается с началом вегетации. Это эндогенное содержание ингибиторов непосредственно связано с морозостойкостью отдельных сортов (Sarkisova с сотр., 1976).
Фитогормоны в репродуктивных органах
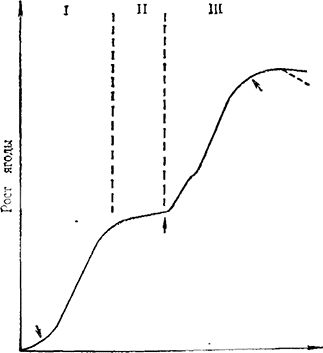
Рис. 1. Динамика содержания ауксинов во время роста ягоды винограда
I — цветение; II — начало созревания; III — физиологическая зрелость
Ауксины. Впервые ауксиноподобные вещества в ягодах винограда установили Nitsсh с сотр. (1960) и Сооrubе (1960). При помощи теста A vena mesocotyle установлено повышение содержания ауксиноподобных веществ в I фазе роста ягод (рис. 1) и понижение его при временном приостановлении роста во II фазе. Исследования Alleweldt и Hifny (1972) и Farmahan и Pandey (1976) подтверждают этот ход ауксиноподобных веществ.
Ауксин в незначительной концентрации доказан в ягодах при помощи газовой хроматографии (Сооmbе, Hale, 1973) и хроматографии с использованием жидкости под большим давлением (During, 1977). Применение гиббереллина калия повысило уже через 3 h после обработки активность ауксинов в ягодах; этот повышенный уровень ауксинов вновь понижается во II фазе роста ягод (Bertrand, Weaver, 1972а). Во время стратификации активность ауксинов в семенах повышается, достигая максимума при прорастании (Сhоhan, Dhillоn, 1976; Касhru с сотр., 1969).
Цитокинины. Новейшие исследования при помощи теста в каллюсе сои показывают, что неидентифицированные до сих пор активные вещества (Waitz, 1975; Сhаско с сотр., 1976; Lilov, Andonova, 1976) появляются в ягодах преимущественно в начале их роста. Относительно содержания цитокининов в отдельные фазы роста имеется немного данных, которые, однако, совпадают. Waitz (1975) установил совсем незначительные концентрации цитокинина в соцветиях в полном цветении, в то время как Lilov и Andonova в этой фазе обнаружили большие концентрации, чем в развивающихся впоследствии ягодах. Пробы в исследованиях брали в один, соответственно в два срока. В разгар цветения и во время образования ягод Lilov и Andonova обнаружили незначительную концентрацию цитокининов, а и Waitz сообщает о цитокининовом минимуме. В противовес этому Сhаско с сотр. (1976) установили максимальные значения, уменьшающиеся в фазу интенсивного роста (I фазу).
Если пренебречь наблюдаемым в III фазе повторным повышением содержания цитокинина, то можно принять, что сделанные констатации о появлении цитокининов в ягодах преимущественно в первую фазу усиленного роста в значительной мере совпадают.
Гиббереллины. Наряду с ауксинами Сооmbе (1960) впервые установил гиббереллиновую активность в незрелых ягодах многих сортов. С началом нарастания ягод Iwahоri с сотр. (1968), Itо с сотр. (1969), а также Lilον и Christov (1977) обнаружили максимальное содержание гиббереллина, задерживающие на одном уровне около двух недель, а затем понижающееся. До сих пор повышение содержания гиббереллиноподобных веществ в III фазе созревания было установлено только Farmahan и Pandey (1976). Бессемянные сорта почти без исключения отличаются меньшей активностью гиббереллина по сравнению с сортами с семенами (Сhailakhуаn, Sarkisova, 1965; Weaver, Pool, 1965а,b; Lilov, Christov, 1977; Nazemi11e, 1977), что выражается несколько замедленным нарастанием незрелых ягод с семенами (Iwahori с сотр., 1968). Незначительное содержание гиббереллина в незрелых ягодах соответствует его меньшим значениям также в листьях, соцветиях и гребнях, тогда как у сортов с семенами в генеративных органах установлено большое содержание гиббереллина, чем в вегетативных (С hailakhyan, Sarkisova, 1965). Weaver и Pool (1965а) обнаружили активность гиббереллиноподобных веществ до оплодотворения в узлах, несущих соцветия. Кольцевание вызывает изменения в спектре гиббереллинов, что означает, что содержание гиббереллина в кислой фракции повысилось, а в нейтральной — понизилось (Weaver, Pool, 1965 b, с).
Абсциссины. О локализации абсциссиновой кислоты в ягодах винограда сообщают Coombe и Hale (1973), Inаbа с сотр. (1976) и During (1973, 1974, 1977), которым при помощи газовой хроматографии, соответственно хроматографии в жидкости под высоким давлением, удалось установить увеличение этого гормона с началом созревания ягод и понижение — перед окончательным созреванием. During (1974) установил почти параллельное повышение абсциссиновой кислоты в мякоти ягод и семенах, в то время как Coombe и Hale (1973) не удалось установить увеличения абсциссиновой кислоты в семенах; Lilov и Angelova (1977b) и Scienza (личные сообщения) также обнаружили лишь незначительное количество абсциссиновой кислоты в семенах. Lott (1968, 1969), однако, установил в выделенных семенах винограда высокое содержание абсциссиновой кислоты, которое уменьшается во время стратификации параллельно повышению степени их готовности к прорастанию. Констатации (неопубликованные) During и данные Chоhan и Dhil1 on (1976) подтверждают эту зависимость. По мнению Coombe (1976a,b) содержание абсциссиновой кислоты повышается примерно в течение 10 недель после цветения, причем в кожице ягод увеличение наступает раньше и быстрее, чем в мякоти.
Этилен. Подобно неклимактерическим плодам черешни (не претерпевающим резких изменений в дыхании во время созревания), в ягодах винограда были установлены совсем незначительные концентрации этилена, во время созревания проявляющие тенденцию к снижению (Coombe, Наlе, 1973). В новейших исследованиях Inаbа с сотр. (1976), Косh (1977) и Аllеwеldt и Koch (1977) также установлено внутриклеточное содержание этилена. Независимо от относительного значения (на ягоду или грамм свежего веса), в начале II фазы установлено повышение содержания этилена, которое, однако, перед началом созревания вновь быстро понижается.