К. Д. Стоев
СТЕБЛЕВЫЕ ЧАСТИ
РОСТ ПОБЕГОВ

Рис. 1. Динамика роста междоузлий
Кружочками обозначены междоузлия между двумя усиками (симподиальная часть побега). По абсциссе — номер междоузлия от основания побега
Рост побегов виноградной лозы изучался многими авторами еще в конце прошлого и начале настоящего столетия, и немало вопросов, касающихся механизма роста, а также зоны наиболее интенсивного роста, уже полностью выяснены. Выводы из этих исследований нашли отражение в руководствах по виноградарству, поэтому на них останавливаться мы не будем.
Здесь отметим только, что удлинение побега происходит в основном за счет растяжения клеток у основания 2-го, 3-го междоузлия, считая сверху, иными словами, осуществляется т. наз. интеркалярный рост (Gаrd, 1903; Мержаниан 1939, 1951, 1967). Эмбриональный рост за счет деления клеток верхушечной меристемы на кончике побега, по мнению Мержаниана (1939), дает очень незначительное удлинение побега.
По данным Тодорова (1970), максимальный прирост междоузлия обнаруживается, когда оно становится 5-6-м, считая сверху, и заканчивается, когда оно становится 8—9-м.
Исследования Мержаниана (1948) показали, что рост междоузлий неодинаков — у междоузлий, заключенных между узлами с усиками (симподиальное строение), он проходит интенсивнее, чем у соседних междоузлий, имеющих моноподиальное развитие (рис. 1). Позже Jaquinet и Simon (1977) установили, что после 3 — 4-го узла на побеге следуют циклически одним за другим короткое, среднее и длинное междоузлия. Эта структура роста побега, по мнению авторов, определяется эндогенной ритмикой и тесно связана с расположением гроздей и усиков.
Рост молодого побега начинается после распускания почек и в зависимости от особенностей сорта и условий среды продолжается, как правило, до начала вызревания побегов, а в отдельных случаях и до физиологической зрелости ягоды. По мнению некоторых авторов, максимальная энергия роста побегов обнаруживается в период цветения. Winkler (1962) отмечает, что в начале вегетации рост идет медленно. Затем, по мере повышения средней температуры, рост побегов ускоряется, достигая максимума накануне цветения (большой период роста). В начале цветения бурный рост замедляется вплоть до начала вызревания побегов. Подобный ход роста побегов наблюдали также В е р е ш с сотр. (1977).
Уменьшение темпа роста побегов происходит неравномерно, подобно неравномерному усилению темпа их роста. Поэтому кривая, выражающая динамику роста побегов по данным Winkler (1962), является как бы идеальной одновершинной кривой (рис. 2).
Почти к аналогичным выводам приходит Солдатова (1958). По ее наблюдениям, прирост побегов достигает максимальной величины перед цветением. Матвеев и Коваль (1975) также считают, что максимум роста побегов достигается накануне цветения. По данным же Мержаниана (1939, 1951, 1967), наиболее интенсивен рост побегов в фазе цветения. В связи с этим Мержаниан отмечает, что вершина кривой роста мало меняет свое положение по отношению к фазам вегетации лозы и всегда будет находиться ближе к фазе цветения.
Несколько иными являются сведения о динамике роста побегов в условиях более низких температур во время прохождения фазы цветения.

Рис. 2. Суточный рост побегов (в десятых долях дюйма.)
Poenaru c сотр. (1961) установили, что в большинстве случаев максимальный прирост побегов достигается после цветения и реже — во время цветения. Они не констатировали ни одного случая, когда максимальный прирост побегов обнаруживался бы до наступления периода цветения.
Ников (1962), изучавший рост побегов ряда сортов, установил, что не имеется строгой связи между цветением и периодом наиболее интенсивного роста. Иногда максимальный рост совпадает с наступлением цветения, иногда он предшествует ему или же наступает по окончании цветения (табл. 1).
Из приведенных данных видно, что срок наступления цветения и период максимального прироста меняются по годам.
Таблица 1
О сроках наступления фазы цветения и максимального прироста побегов
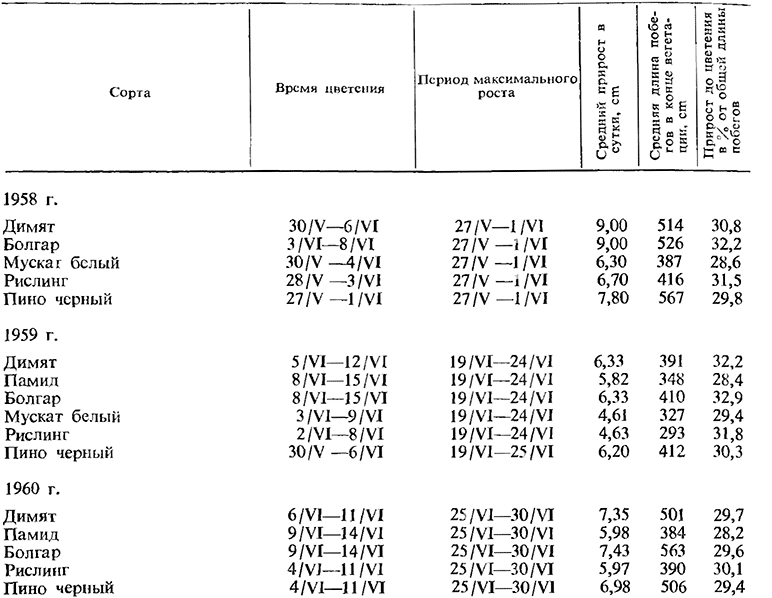
Несмотря на то, что в условиях одного и того же года цветение не наступает одновременно для различных сортов, максимальный рост побегов обнаруживается всегда одновременно. Отсюда можно сделать вывод, что максимальный рост не зависит от биологических особенностей сортов, а обусловливается метеорологическими условиями, и, в первую очередь, температурным фактором. В условиях более высоких среднемесячных майских и июньских температур 1958 г. (в среднем на 1 — 3°С выше) максимальный рост почти полностью совпадал с наступлением периода цветения или же немного опережал его. В годы с более низким температурным напряжением (1959 и 1960) максимальный рост устанавливался через 4—14 дней после окончания цветения. Из этого можно заключить, что оптимальная температура цветения и роста побегов не совпадает. В силу этого совпадения периода цветения с максимальным приростом побегов можно ожидать только тогда, когда оптимальная для роста побегов температура наступает до цветения или к началу периода цветения.
Рост побегов имеет также суточную динамику. В литературе по общей физиологии растений (Максимов, 1958) отмечается, что рост растений ночью интенсивнее, несмотря на пониженную температуру. Исследования роста побегов виноградной лозы в дневные и ночные часы не подтверждает полностью это положение. Так, но данным Солдатовой (1958), в период начала и максимального роста побегов (фаза цветения) наибольший рост отмечался между 12 и 16 ч. По ночам, с 20 до 8 ч., когда температура воздуха снижалась до 5—8°С, рост затухал, а с 8 до 20 ч. снова повышался (рис. 3).

Рис. 3. Интенсивность роста побегов (mm) в течение суток
1 — с 8 до 12 ч.;2 — с 12 до 16 ч.;3 — с 16 до 20 ч.; 4 — с 20 ч.; 5 — с 24 до 4 ч.; 6 — с 4 до 8 ч.; 7 — температура
Исследования Никова (1962) в основном подтверждают выводы Солдатовой, что в дневные часы он более интенсивен, причем сортовых различий не имеется. В отличие от Солдатовой, однако, Ников установил, что дневной прирост значительно превалирует над ночным* в начальные этапы роста (май) — около 65 % от общей величины суточного прироста. В июне дневной и суточный прирост почти выравниваются (50%), однако к концу месяца уже отмечается тенденция к ослаблению дневного прироста. Во второй половине июля дневной прирост составляет лишь 1/4 до 1/3 суточного прироста (табл. 2).
Таблица 2.
Соотношение дневного и ночного прироста побегов (%) в течение вегетации (1959 г.) | ||||
Сорт | 16—18/V | 5—7/VI | 28—30/VI | 18—20/ VII |
Димят | 67,4 | 54,4 | 49,5 | 34,0 |
Памид | 64,8 | 53,3 | 47,5 | 28,2 |
Болгар | 68,8 | 56,1 | 51,3 | 35,0 |
Рислинг | 62,5 | 52,4 | 46,7 | 25,5 |
Таким образом соотношение между дневным и ночным приростом — величина непостоянная и меняется по фазам вегетации, главным образом под влиянием температуры. В начальные этапы вегетации (до цветения) температура ночью значительно ниже оптимальной, поэтому она подавляет рост. К концу роста дневная температура значительно выше оптимальной, вследствие чего теперь она является сдерживающим рост фактором.
* Прирост измерялся в 6 и 18 ч.
Рис. 4. Сопоставление сезонных изменений прироста побегов (cm) и температуры воздуха
1 — температура; 2 — прирост побегов; 3 — число междоузлий
Значение температурного фактора для роста побегов выявляется также при сопоставлении сезонных изменений прироста и температуры. Из исследований Никова (1962) видно, что суточный прирост побегов идет почти параллельно величине суточной температуры, что указывает на их функциональную зависимость (рис. 4). Взаимосвязь настолько тесна, что любому повышению или снижению температуры соответствует почти адекватное повышение или снижение интенсивности роста.
Jaquinet и Simon (1977) показали, что среднее удлинение побегов совсем точно совпадает с кривой средней величины температуры соответствующего периода (рис. 5).
Согласно данным Dufrenоу (1932), удлинение побегов за каждый 24-часовой период (с 9 ч. утра до 9 ч. следующего дня) находилось в прямой зависимости от величины температуры предшествующих суток. Таким образом, температура предшествующего дня как бы индуцирует величину прироста следующего дня. Взаимосвязь, однако, устанавливается лишь в период интенсивного роста (в мае и июне). С ослаблением темпа роста реакция на повышение температурного фактора непрерывно уменьшается, а поэтому и кривые сезонных изменений прироста побегов и температуры расходятся (рис. 4).
Аналогичную зависимость между суточной температурой и приростом побегов установил также Bouard (1966). Константинеску и Ошлобяну (1956), изучавшие динамику прироста сорта Коарна Нягра, сделали неправильный вывод, что рост во время цветения снижается и что это функционально связано с прохождением процесса цветения и оплодотворения. На самом деле снижение темпа роста находится в прямой зависимости от величины суточной температуры, которая снизилась как раз во время цветения.
Влияние остальных факторов среды (осадков, атмосферной влажности и солнечной радиации) на рост побегов не очень заметно. По-видимому, доминирующее влияние из метеорологических факторов оказывает температура. По данным Стоева и Занкова (1962), продолжительность дневного освещения сказывается подавляюще на приросте побегов как только длина дня уменьшится до 8—10 h.

Рис. 5. Взаимосвязь между удлинением побегов и температурой
1 — среднесуточный прирост (mm); 2 — среднесуточная температура (°С)
На рост побегов оказывает влияние также ряд других факторов. Как известно, близость побега к корневой системе обусловливает большую энергию роста. В связи с этим побеги, выросшие из сучка замещения или же многолетней части, всегда растут интенсивнее, чем побеги на стрелке (Стоев и Рангелов, 1969). Еще ниже темп роста и величина прироста побегов, развившихся на высоком штамбе. Побеги, развившиеся из различных почек по длине стрелки, также растут неравномерно. Как правило, рост проходит интенсивнее у побегов, находящихся у основания и в конце стрелки. Здесь оказывает влияние полярность. В связи с этим Веssis (1977) отмечает, что акротония (полярность) виноградной лозы является результатом двух особенностей: с одной стороны, побеги на конце стрелки отличаются более сильным ростом, чем у ее основания, а, с другой, процент проросших почек снижается к основанию стрелки. По данным Стоева и Рангелова, в динамике роста побегов на плодовой стрелке сорта Мискет червен обнаружено два максимума — первый в секторе 3 — 4-го глазка, второй — в секторе 10—12-го глазка стрелки. Для роста побегов имеет значение также способ подвязки стрелки (Неделчев и Тодоров, 1966). Исследования Тодорова (1977) показали, что способ подвязки может вызвать существенные изменения среднего веса гроздей, урожая в пересчете на один глазок, среднего веса и сахаристости ягод.
В естественных условиях рост кустов зависит от коррелятивного подавления расположенных ниже побегов, в то время как прорастание почек вначале зависит от их особенностей, которые дают им возможность развиваться раньше или позже (Веssis, 1977). Если какая-либо почка начнет развиваться позже смежных с ней, в некоторых случаях она не сможет прорасти вследствие коррелятивного подавления. Skene (1972) установил, что кольцевание приводит к некоторому уменьшению длины побега, однако их сухой вес не уменьшается. Кольцевание вызывает также значительное уменьшение активности цитокининов (фракция А). Это уменьшение не возрастает после кольцевания.

Рис. 6. Влияние толщины стрелки (mm) на длину побегов (cm)
1 — сорт Болгар; 2 — сорт Димят; 3 — сорт Мискет червен
Из условий выращивания изучалось влияние нагрузки глазками, орошения, удобрения, обрезки зеленых частей, сроков обрезки и пр. на рост побегов. Влияние некоторых из этих факторов рассматривается в соответствующих разделах. Отметим лишь, что имеющиеся сведения в этом направлении нередко разноречивы. При этом чаше всего изучалась не динамики роста, а общий прирост по весовым показателям или же по общей длине прироста.
Тодоров (1978) показал, что толщина стрелки, из которой развиваются побеги, заметно сказывается на их росте. Из данных рис. 6 видно, что с увеличением толщины стрелки с 6-8 до 8—10, 10—12, 12 — 14 и 14—16 mm удлиняются и побеги, выросшие из их почек. При этом корреляция не имеет прямолинейного характера — после достижения определенной оптимальной толщины стрелки средняя длина побегов начинает уменьшаться. Что же касается общего прироста побегов, то он увеличивается почти прямолинейно в зависимости от толщины стрелки, в особенности у сортов Болгар и Мискет червен.
По данным Bouard (1966), срок обрезки не сказывается существенно на росте побегов и длине междоузлий. Удобрение также не оказало заметного влияния на прирост побегов.
Исследования Димитрова (неопубл. данные) показали, что подвои Рупестрис дю Ло и Берландиери х Рипария Кобера 5ББ оказывали большее влияние на темп роста побегов привоя и их общий прирост, чем подвой Шасла х Берландиери 41 Б. При этом влияние подвоя на рост побегов привоя сильнее выражено у сорта Уньи белый, чем у сорта Каберне Совиньон.
Стоев и Рангелов (1969) установили также, что для роста побегов существенное значение имеет корневая система. Как правило, наиболее интенсивный рост побегов устанавливался при прививке сорта Мискет червен на подвой Рупестрис дю Ло, наименее интенсивный — при выращивании сорта на собственном корню. Среднее положение по энергии роста занимали побеги, развивавшиеся на подвое Шасла х Берландиери 41Б (рис. 7). При этом побеги, развитые на корневой системе Шасла х Берландиери 41Б, имели в среднем 27 междоузлий, в то время как побеги, выросшие на корневой системе подвоя Рупестрис дю Ло, имели в среднем 20 междоузлий, а на собственном корню— 17. Это показывает, что корневая система влияет и на среднюю длину междоузлия (рис. 8).

Рис. 7. Рост побегов сорта Мискет червен (V. vinifera) в зависимости от корневой системы (cm), среднесуточной температуры (°С) и величины осадков (mm)
1 — на собственном корню (без прививки); 2 — на подвое Шаслах Берландиери 41 Б; 3 — на подвое Рупестрис дю Ло; 4 — температура

Рис. 8. Влияние корневой системы на длину побегов и число междоузлий на побеге
а — Мискет червен на собственных корнях; б — Мискет червен на подвое ШаслаХ Берландиери 41 Б; в — Мискет червен на подвое Рупестрис дю Ло
Влияние подвоя на рост побегов привоя выражается и в том, что чем сильнее корневая система, тем больше максимальный рост побегов сдвигается на ранние этапы вегетации. В условиях нормального температурного напряжения кусты, привитые на сильнорослый подвой (Рупестрис дю Ло), отличаются, как правило, максимальным ростом побегов до начала цветения.
Максимум роста побегов на подвое Шасла х Берландиери 41Б совпадает с фазой цветения, а на собственном корню наступает после цветения.
Более существенные различия обнаружены в динамике роста и общего прироста побегов различных подвоев. Наблюдения Димитрова (неопубл.) показали, что рост побегов подвоев Кобера 5 ББ, SO4 и др. значительно интенсивнее роста побегов подвоя Шасла х Берландиери 41Б и их общая длина примерно на 1,5 раза больше. Ергесян (1952) обнаружил более интенсивный рост побегов на подвое Кобера 5 ББ по сравнению с 3309 и 101-14.
Negreanu с сотр. (1977) установили, что опрыскивание L-нафтилуксусной кислотой, 2,4 ДФУ, гиббереллином и фторамидом стимулирует рост побегов, а ССС (хлорхолинхлорид) подавляет его. Alleweldt (1977) также показал, что гиббереллиновая кислота стимулирует рост побегов в условиях длинного и короткого дня, ССС, наоборот, сильно подавляет рост.
Подавление роста побегов под влиянием ССС установили Куртеева, Колесник (1972) и Куртеева (1977), причем авторы наблюдали и другие явления — утолщение луба, увеличение в нем лубяных волокон и усиленное одревеснение. Farcioli с сотр. (1971) показали, что реакция сортов на ССС специфическая, однако у всех обработанных сортов усиливается пасынкообразование и увеличивается урожай.
На росте побегов подвойных лоз сказывается также система выращивания. Эти вопросы, однако, недостаточно изучены. Частично они рассматриваются в третьем томе этого труда.
Недостаточно изучены и вопросы, касающиеся роста пасынков. По данным Раваза (цит. по Мержаниану, 1939), рост пасынка у самого нижнего узла главного побега незначителен, затем у 3 — 5-го узла он достигает максимума, а на следующих узлах величина пасынков все больше и больше уменьшается.
По наблюдениям К. Стоева и 3. Занкова (неопубл. данные), пасынки на побегах сортов Зарчин и Мискет червен, выросших из почек сучка замещения, растут сильнее, чем пасынки на побегах, развитых на стрелке. Дальнейшие исследования Стоева и Рангелова (1969) подтвердили это положение и показали, что рост пасынков зависит также от корневой системы. Наиболее длинные пасынки установлены на побегах (сорт Мискет червен), развивающихся на подвое Рупестрис дю Ло, меньшие — на подвое Шасла х Берландиери 41Б и наиболее коротки пасынки на побегах, растущих на корнесобственной системе. Характерно еще то, что наиболее развитые пасынки на побегах, растущих на подвое Рупестрис дю Ло, находятся в нижней половине побега, в то время как они достигают более сильного развития в верхней половине побега у кустов без прививки.
По данным Тодорова (1977), пасынки растут сильнее, если против них нет усика. Длина пасынков больше, если они вырастают из узла без усика. Установлено также, что пасынкообразовательная способность разных сортов неодинакова (Ульяницкая, 1975). В дальнейшем Тодоров (1978) убедительно показывает, что общая длина побегов пасынков сильно зависит от толщины основных побегов (r=0,974—0,995), причем общая длина пасынков складывается главным образом из их индивидуального роста.