Введение
Сложные эфиры, образующиеся в результате действия кислот на спирты, обязательно присутствуют в винах. Некоторые эфиры находятся в небольших концентрациях в винограде. Другие, главным образом нейтральные сложные эфиры, представляют результат жизнедеятельности дрожжей и бактерий (этилацетат, эпиллактат и др.). Во время созревания вина кислоты и спирты находятся в состоянии медленной реакции друг с другом и образуют главным образом кислые сложные эфиры, нелетучие в нормальных условиях дистилляции (этилтартрат, этил-сукцинат и др.). Реакции этерификации протекают медленно и обычно ограничены. Они становятся ощутимыми в винах только по истечении многих месяцев. Даже через полвека выдержки они не достигают своего предела старения.
Запах сложных эфиров одноосновных жирных кислот, высших по отношению к уксусной кислоте, не сравним с ароматами и букетами, которые могут встречаться в винах. С другой стороны, содержание сложных эфиров в старых винах не имеет никакой связи с их качеством. Оно является в действительности результатом процессов, общих для всех вин, тонких и ординарных. С давних пор известно, что вина, самые богатые сложными эфирами, часто оказываются самыми ординарными, имеющими больше всего пороков и хуже всех сохраняющимися.
Другая серия наблюдений привела к аналогичным выводам. С началом брожения сусло теряет присущий ему запах виноградного сока и приобретает характерный аромат молодого вина. Этот аромат, который можно собрать, барботируя через спиртовой раствор выделившийся при брожении углекислый газ, не связан со сложными эфирами. Барботажная жидкость, аромат которой довольно аналогичен аромату самого вина, не имеет заметного количества сложных эфиров. Согласно работам Кордонье речь идет о терпеновых углеводородах или соединениях ароматического ряда. Хроматография в газовой фазе способствовала идентификации этих веществ.
Закономерности образования сложных эфиров
Было показано, что этерификация (реакция между кислотами и спиртами) бывает медленной и ограниченной. Если, например, смешать раствор уксусной кислоты с раствором этилового спирта, то раствор постепенно становится менее кислым в результате нейтрализации кислоты спиртом. Происходит образование нейтральных веществ: сложного эфира уксусной кислоты или этилацетата и воды:
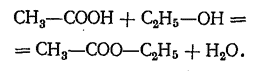
Эту реакцию приравнивали к нейтрализации кислоты щелочами. Однако имеются фундаментальные различия. Насыщение кислот основаниями происходит мгновенно; это—реакция ионов, т. е. процесс распада присутствующих веществ. В противоположность этому этерификация происходит очень медленно, и ее скорость в значительной мере зависит от температуры. Этерификация, которая при обычной температуре длилась бы годы, при температуре 100°С завершается за несколько дней, а при температуре 200°С — за несколько часов. Это — мономолекулярная реакция; здесь выступают отдельные, недиссоциированные молекулы кислоты.
Если проследить образование этил-ацетата в вышеупомянутой смеси (уксусная кислота + этиловый спирт) в зависимости от времени, то констатируют, по мере того как протекает этерификация, все более и более выраженное замедление реакции, которая в конце концов прекращается. К этому моменту еще не вся кислота нейтрализована спиртом: 1/3 уксусной кислоты и спирта остается свободной, а 2/3 превратились в этилацетат. Реакция достигла своего конечного предела; система находится в состоянии равновесия.
Если смешать растворы этилацетата и воды, что, в сущности, означает то же самое, что осуществить полную реакцию кислоты и спирта, такая смесь не будет в состоянии стабильного равновесия и будет вести себя как смесь уксусной кислоты и спирта. Будучи сначала нейтральной, она постепенно становится кислой. В ней происходит гидролиз, обратный реакции этерификации. Как и этерификация, эта реакция протекает очень медленно, и ее скорость в значительной мере зависит от температуры. По мере развития она все более замедляется. Реакция останавливается задолго до того, как разложится весь этилацетат. Состояние равновесия раствора характеризуется такими же пропорциями тех же веществ, как и в случае эквивалентной смеси спирт+кислота. Следовательно, происходят две реакции, которые себя взаимно ограничивают, — этерификация и омыление. Вышеприведенную реакцию действия кислот на спирты можно переписать со знаком обратимости (двойная стрелка).
Многоосновные кислоты или поликислоты в зависимости от того, участвуют ли в реакции со спиртом одна или несколько их карбоксильных групп, образуют кислые сложные эфиры или нейтральные сложные эфиры почти так же, как они образуют кислые или средние соли. Следовательно, винная кислота, поскольку и она способна образовывать кислый битартрат и виннокислый калий, может дать два сложных эфира: сложный моноэфир или кислый этилтартрат:


Состояние равновесия конечной системы зависит от концентрации каждого из компонентов. Оно регулируется законом действующих масс. Если обозначить через А, О, Е, [H2O] концентрации, эквивалентные кислоте, спирту, сложному эфиру и воде, в равновесной системе, то можно записать
К не зависит от температуры и природы органических кислот. Значение К для образования этиловых эфиров близко к 4. Когда ищут пропорцию сложных эфиров в равновесной системе, предыдущее отношение можно записать так:

В табл. 14.1, рассчитанной с использованием закона действующих масс и эмпирической формулы Вертело, дано процентное отношение свободной кислотности, способной преобразовываться в сложные эфиры в зависимости от содержания спирта. Причем предполагается, что вина имеют 20 г/л сухого экстракта.
Таблица 14.1
Количество свободных кислот (в %), превращаемых в сложные эфиры, в зависимости от спиртуозности среды
| Содержание спирта, % об. | По закону действующих масс | По формуле |
8 | 10,0 | 9,4 |
9 | 11,2 | 10,2 |
10 | 12,4 | 11,0 |
11 | 13,6 | 11,7 |
12 | 14,8 | 12,5 |
13 | 16,0 | 13,3 |
14 | 17,2 | 14,0 |
15 | 18,4 | 14.8 |
Образование эфиров в простых растворах
В табл. 14.2 дано при двух pH и содержании спирта 10% об. количество этерифицированных свободных кислот при нагревании до 100°С через 24 ч и через 30 дней. В первом случае можно сравнивать скорости образования сложных эфиров в зависимости от природы кислоты, во втором — получают примерное представление о пределе реакции.
Таблица 14.2
Количество свободных кислот (в %), превращенных в сложные эфиры, в зависимости от pH
Кислота | pH 3 | pH 4 | ||
24 ч, | 30 дней, | 24 ч. | 30 дней, 100° С | |
Янтарная | 8,4 | 10,2 | 3,9 | 9,3 |
Яблочная | 9,0 | 10,2 | 3,8 | 9,1 |
Молочная | 8,5 | 9,8 | 3,0 | 8,8 |
Винная | 5,0 | 9,3 | 1,5 | 8,6 |
Лимонная | 4,4 | 6,7 | 3,0 | 6,3 |
Уксусная | 2,7 | 8,7 | 0,8 | 7,5 |
Пропионовая | 2,4 | 9,0 | 1,2 | 7,7 |
Масляная | 1,4 | 8,7 | 0,7 | 7,6 |
Из табл. 14.2 видно, что природа кислоты и pH среды представляют собой факторы, определяющие скорость реакции. Ионы Н+ являются очень активными катализаторами реакций образования сложных эфиров. Предельный коэффициент этерификации, вычисленный по закону действующих масс, равен 12,4% от первоначальной кислотности и по формуле Вертело равен 11%. Эти значения не были достигнуты к концу месяца нагревания до 100°С при pH 3 ни одной из кислот, а также после 8 дней нагревания при 100°С и pH 1,8.
Природа эфиров, образованных многоосновными кислотами, зависит от pH. Соотношение нейтральных эфиров будет тем больше, чем ниже pH. Для значений pH, которые бывают в винах (2,8—3,8), эта концентрация очень мала, и можно предполагать, что вина не содержат нейтральных этиловых эфиров винной, яблочной, лимонной и янтарной кислот.
Образование эфиров в винах
Сложные эфиры вина образуются или путем химической реакции во время созревания, или же биологическим путем — под действием дрожжей во время брожения или под влиянием бактерий. В целом оба эти процесса в одинаковых пропорциях ответственны за происхождение сложных эфиров вина. В результате химической этерификации образуются прежде всего кислые сложные эфиры, а вследствие биологической этерификации — главным образом нейтральные сложные эфиры.
Содержание общих эфиров вина определяется, его составом и возрастом. Оно колеблется от 2—3 мг-экв/л в молодых винах до 9—10 мг-экв/л в старых винах. Его увеличение особенно заметно в течение первых двух лет хранения. В дальнейшем это возрастание замедляется. Образовавшееся количество всегда остается далеким от предела. Вина, выдерживавшиеся около 50 лет, содержат всего лишь 75% дозы, соответствующей концу реакции. Отношение дозы общих эфиров в данный момент к предельной дозе может указывать очень приближенно на возраст вина (табл. 14.3).
Нейтральные сложные эфиры винной, яблочной и лимонной кислот редко достигают концентрации 0,75 мг-экв/л, и, вероятно, их образование активизируется только химическими катализаторами. Эти сложные эфиры образуются при длительной выдержке вина, в молодых винах не встречаются даже их следы. Этиллактат находится в довольно больших количествах в винах, богатых молочной кислотой. Он появляется во время спиртового и яблочно-молочного брожения.
Таблица 14.3
Отношение дозы общих сложных эфиров к предельной дозе в зависимости от возраста вина
| Возраст вин | Крайние значения отношения | Средние значения отношения |
От 22 до 43 лет | От 0,73 до 0,79 | 0,75 |
» 6 » 10 » | » 0,57 | 0,66 |
» 4 » 5 » | » 0,59 | 0,64 |
3 года | » 0,49 | 0,62 |
2 » | » 0,50 | 0,56 |
8 мес | » 0,28 | 0,34 |
В старых винах кислые сложные эфиры бывают в концентрации нескольких мг-экв/л. Среди них этилтартрат может составлять 1,5 мг-экв/л и больше, но этерификация винной кислоты остается всегда довольно далекой от предела равновесия. Статика образования сложных эфиров допускает присутствие лишь незначительных доз этил-фосфатов и глицерофосфатов.
Следует всегда иметь в виду, что образование сложных эфиров у органических кислот вина происходит индивидуально и независимо друг от друга и что оно никогда не достигает теоретического предела ни для одной из них. Для того чтобы получить заметное повышение содержания сложных эфиров в винах, требуется длительная выдержка при температурах, более высоких, чем те, которые могут вынести вина, не претерпевая денатурации. Пастеризация не может увеличить содержание сложных эфиров.
Образование этилацетата уксусными бактериями
Установлено, что этилацетат, особенно образованный уксусными бактериями, а не уксусная кислота, как считали долгое время, представляет собой вещество, ответственное за истинные признаки уксусного скисания, ощутимые обонянием. Его образование вначале имеет чисто биологический характер, и медленные химические реакции созревания на этом этапе не проявляются. Этилацетат вырабатывают дрожжи во время брожения и в некоторых случаях — уксуснокислые бактерии.
Содержание этилацетата, встречающееся в нормальных, хотя и не абсолютно здоровых винах, может изменяться, например, от 40 до 160 мг/л. Чистые культуры эллиптических дрожжей всегда дают небольшое количество сложных эфиров. Ниже будет показано, что некоторые расы дрожжей обладают особо выраженными свойствами и образуют большое количество этил-ацетата. Но прежде всего это относится к уксуснокислым бактериям или Aceto-bacter, которые образуют во время окисления этилового спирта вин в уксусную кислоту более или менее значительные количества этилацетата в зависимости от вида бактерий и условий температуры.
Присутствие в микробиальных клетках специфического фермента (эстеразы) обусловлено биологической этери-фикацией. Коэффициент биологической этерификации не подчиняется закону действующих масс и может даже превзойти предел для этого химического равновесия. Некоторые расы дрожжей Hansenula также образуют небольшое количество уксусной кислоты и много этилацетата. Сложные эфиры образуются внутри клетки и только из кислот, которые там вырабатываются. Отсюда следует, что во время брожения кислоты сусла не образуют сложных эфиров, не этерифицируются; с другой стороны, уксуснокислые бактерии вырабатывают только этилацетат. Это хороший пример специфичности ферментативных реакций: сложный эфир образуется исключительно за счет кислот, которые вырабатывает клетка.
С другой стороны, если создать условия для развития уксуснокислых бактерий на поверхности синтетической питательной среды или вина, то можно наблюдать, что содержание образовавшегося этилацетата возрастает пропорционально образованию уксусной кислоты. Коэффициент этерификации, или фракция кислоты, которая этерифицируется, представляет собой характеристику расы Acetobacter и зависит от температуры. Он колеблется от 2 до 10%, в данном же случае возможный предел равен 11,2%. Acetobacter rancens обладает значительно меньшей способностью образовывать сложные эфиры, чем Acetobacter ascendans и Acetobact. xylinum. Содержание этилацетата выше при 21 С, чем при 11 или 31°С.
Следовательно, этерификация бактериями не имеет ничего общего с этерификацией химической. Чтобы этерифицировать 7% присутствующей уксусной кислоты химическим путем при температуре 100°С, требуется около двух дней. Бактериями этерифицируется 80% уксусной кислоты при 21°С через два дня. Частичная биологическая этерификация уксусной кислоты происходит практически мгновенно по сравнению с химической этерификацией. Кислоты, которые этерифицируются намного легче (молочная, яблочная), но чуждые бактериям, совершенно не этерифицируются при контакте с ними.
Биологическая этерификация обратима; в присутствии этилацетата бактерии гидролизуют его. Именно поэтому в уксусах, где спирт целиком превращается в уксусную кислоту, больше не находят этилацетата, и запах уксуса не похож на запах скисшего вина. С другой стороны, в процессе уксусного скисания с поверхности пленки должно происходить значительное испарение этилацетата, если судить об этом по запаху, появляющемуся с первого момента образования уксуса. Считают, что это испарение может быть также результатом влияния температуры.
В конечном счете образование уксусной кислоты и этилацетата бактериями Acetobacter представляется в виде двух параллельных, но совершенно независимых одно от другого явлений. Это образование связано с двумя очень различными функциями клетки: образованием уксусной кислоты в процессе дыхания и образованием этилацетата при высоком содержании эстеразы в клетке.
Образование этилацетата дрожжами
По своей способности образовывать сложные эфиры винные дрожжи можно подразделить на несколько групп. К первой группе, которая охватывает все дрожжи родов Saccharomyces и Torulopsis stellata, относятся дрожжи со слабовыраженной эфирообразующей способностью. Концентрация образовавшегося этилацетата не превышает 30—50 мг/л при брожении с доступом воздуха и 20—30 мг/л при брожении без доступа воздуха. Такое количество не оказывает влияния на букет вина, во всяком случае, неблагоприятного. Отсюда следует, что дрожжи этой группы можно считать хорошими винодельческими дрожжами. Вторая группа включает несколько видов дрожжей, обладающих немного более высокой способностью вырабатывать этилацетат (от 60 до 110 мг/л при брожении с доступом воздуха и от 40 до 80 мг/л без доступа воздуха), но значительно реже встречающихся на практике: Меschnikowia pulcherima, Hanseniaspora и Brettanomyces.
В противоположность всем вышеперечисленным видам Saccharomycodes Ludwigii образуют в 2 раза больше этилацетата в анаэробиозе, чем на воздухе, но дают меньше уксусной кислоты. Их эфирообразующая способность связана с различным метаболизмом. Продукты, полученные в результате брожения с дрожжами Saccharomycodes ludwigii, характеризуются четко выраженным запахом этого сложного эфира. Hanseniaspora и Kloeckera также вырабатывают большое количество этилацетата. В результате брожения на чистой культуре дрожжей апикулятус получают вина, у которых при дегустации обнаруживаются признаки уксусного скисания. Поэтому вполне вероятно, что содержание этилацетата в молодых винах во многом зависит от участия этих дрожжей в начале брожения.
Наконец, пленчатые дрожжи Pichia и Hansenula хорошо известны своей способностью вырабатывать сложные эфиры. Наиболее высокие концентрации (примерно 900 мг/л) создают дрожжи Hansenula. Дрожжи Pichia обычно дают менее высокие и не всегда постоянные результаты.
Можно считать, что этилацетат образуется во время спиртового брожения двумя различными путями: а) путем ферментативной этерификации, поскольку общепризнано присутствие эстеразы в дрожжах. Такой процесс, наблюдаемый у Saccharomyces, дает не более 2% всей образуемой уксусной кислоты; б) посредством прямого биосинтеза, связанного у некоторых дрожжей с дыханием, но независимого от респирации у Saccharomycodes. Вполне вероятно, что эти различные пути образования этилацетата у многих видов дрожжей переплетаются.