Онтогенез растений складывается из вегетативного роста и генеративного развития с параллельным прохождением одногодичного органогенеза. У многолетних растений развитие включает и период покоя.
В процессе индивидуального развития в различных тканях органов растения проходят многочисленные физиологические и биохимические процессы и превращения в обмене веществ, специфические для отдельных тканей, органов и растения в целом. С изменением условий среды, возраста и самого растения и отдельных его органов происходят в различные этапы их развития физиологические и биохимические изменения в обмене веществ, сопровождаемые процессом дыхания. Поэтому целесообразно изучение и знание процесса дыхания отдельных органов растения в различные периоды их развития и в онтогенезе в зависимости от условий среды или под влиянием тех или иных агроприемов.
В настоящее время имеется относительно большое число исследований по дыханию органов виноградного растения в онтогенезе для различных условий произрастания и сортов разного происхождения.
Листья
По дыханию листьев представляет интерес работа Сисакяна и др (1948) (табл. 1).
Таблица 3
Дыхание листьев I и II формаций у различных сортов винограда (mg СО2/h на 50 cm3) в 1948 г. (по Сисакяну)
| Фаза | Воскеат | Чилар | Мохали | Гарандмак | ||||
I | II | I | II | I | II | I | II | |
До цветения | 2,5 |
| 1,2 |
| 3,7 |
| 1,2 |
|
Цветение | 2,2 | — | 1,5 | — | 1,5 | — | 1,5 | — |
От конца цветения до начала созревания | 2,3 | 6,0 | 2,1 | 7,0 | 1,5 | 5,5 | 2,1 | 5,7 |
Начало созревания | 4,5 | 5,5 | 4,7 | 6,0 | 1,7 | 3,0 | 1,5 | 3,5 |
Физиологическая зрелость | 1,7 | 5,0 | 1,5 | 2,5 | 2,5 | 4,0 | 2,5 | 3,7 |
Листопад | 1,0 | 1,5 | 0,2 | 1,2 | 1,2 | 1,2 |
| — |
Активность дыхания листьев винограда к листопаду снижается. Дыхание листьев второй формации (более молодых) у всех сортов в несколько раз (до трех и более) превышает активность дыхания листьев первой формации (большего возраста). Максимум активности дыхания листьев второй формации приходится на период после цветения, в то время как у более взрослых (первой формации) максимум наблюдается в начале созревания (сорта Чилар и Воскеат). Как сортовое различие следует отметить, что у сортов Мсхали и Гарандмак максимум дыхания листьев первой формации сдвинут на более поздний период, что можно объяснить более длинным вегетационным периодом этих двух сортов.
Авторы отмечают, что не всегда имеется прямая зависимость между изменениями в интенсивности дыхания и активности ферментных систем. Эти изменения могут быть взаимосвязаны, но не обязательно в прямой зависимости. В свете развиваемых Михлиным и Колесником (1947) представлений обнаруженные различия в активности окислительно-восстановительных ферментов и биоактивных соединений можно объяснить наличием у винограда различных дыхательных систем, причем в различные фазы развития превалирует та или иная система дыхания. Авторы отмечают, что в листьях винограда и яблони, что установлено Сисакяном и Рубиным (1944), полифенолоксидазная система начинает действовать более активно лишь в поздние фазы развития, как и пероксидазная система.
Изменения активности дыхания и работы ферментов слабее выражены для различных сортов по сравнению с условиями среды и возрастом того или иного органа.
В условиях северной лесостепной зоны Молдавии Найденов (1969) провел изучение интенсивности дыхания разновозрастных листьев на терминальных побегах сортов винограда Алиготе и Шасла белая. В исследованиях использован метод Сакса с раздельным определением дыхания и оттока. Результаты проведенной работы показали, что в начале созревания ягод (10— 12/VIII) наиболее высокая интенсивность дыхания отмечалась у верхних листьев (3—4-й сверху). Ко времени прекращения роста побегов она была в 2,5—3,0 раза выше, чем у нижних (табл. 2). Второе место по выделению СO2 на единицу площади листа занимают листья средней части побега, а также листья пасынковых побегов. Наиболее слабая интенсивность дыхания наблюдалась у нижних листьев, которые к этому периоду постепенно снижают свою фотосинтетическую активность и в большей степени участвуют в процессах вызревания лозы, в транспорте и накоплении пластических веществ в многолетней кроне куста и в корневой системе.
Таблица 2
Энергия дыхания листьев разных ярусов (северная лесостепная зона) (10—12/VIII 1958 г.)
Сорт | Расположение листьев на побеге | Выделено СO2, mg/h | |
на 100 g | на dm2 листьев | ||
Алиготе | верхние (3—4 сверху) | 141,2 | 2,70 |
средние | 69,6 | 1,75 | |
нижние (2—4 снизу) | 46,0 | 1,03 | |
пасынковые (1—2 снизу) | 84,0 | 1,68 | |
Шасле белая | верхние (3—4 сверху) | 112,7 | 2,32 |
средние | 60,8 | 1,43 | |
нижние (2—4 снизу) | 42,3 | 0,83 | |
пасынковые (1—2 снизу) | 73,7 | 1,39 | |
В ходе физиолого-биохимических процессов живой клетки в процессе онтогенеза условия агротехники могут вызвать значительные изменения. Определенный интерес в связи с этим имеют исследования Библиной (1956), проведенные в Молдавии на сортах Сенсо и Шасла, по изучению влияния нагрузки побегами куста на интенсивность дыхания листьев винограда при разных площадях питания кустов.
Результаты исследований показали изменения интенсивности дыхания листьев по фазам вегетации, причем наиболее высокая интенсивность дыхания относится к периоду массового цветения, к началу созревания она снижается и несколько повышается в процессе созревания ягод.
Автор отмечает, что в течение всего вегетационного периода дыхание листьев у сорта Шасла выше, чем у Сенсо, и у этого сорта значительно слабее выражены различия между отдельными вариантами площади питания (1,25 х 2,25 и 2,25 х 2,25 т) и нагрузки побегами на куст (20 и 49 побегов/куст) (рис. 2).
В течение вегетации активность дыхания изменяется по фазам и является различной для изучаемых вариантов нагрузки и площади питания.
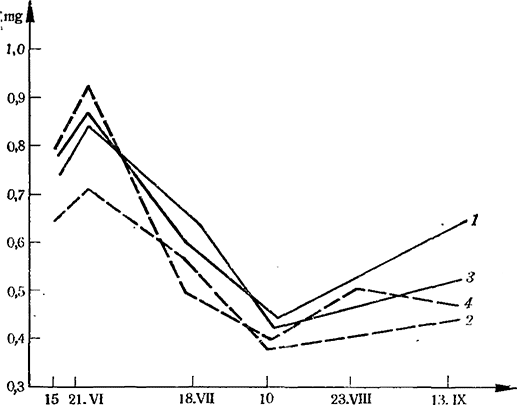
Рис. 2. Интенсивность дыхания (mg СO2) листьев сорта Шасла. Площадь питания 1,25x2,25 и 2,25x2,25
1 — 20 побегов на куст; 2 — 49 побегов на куст; 3 — 24 побега на куст; 4 — 61 побег на куст
В общем перегрузка побегами ослабляет дыхание, особенно при меньшей площади питания, а активность каталазы при этом как показатель жизнедеятельности растения в большинстве случаев усиливается в обоих вариантах площади питания. Ход изменений активности каталазы листьев часто не совпадает с ходом изменений дыхания в том или ином варианте исследований. За весь период вегетации у сорта Шасла активность каталазы выше, чем у Сенсо.
В работе Библиной (1963) проведено изучение изменений содержания аскорбиновой кислоты как фактора, регулирующего превращения сахарозы в окислительно-восстановительных процессах и в азотном метаболизме растительной клетки (Рубин, Арциховская, 1937; Михлин, 1936), а также изучение дыхания. Аскорбиновая кислота принимает участие в фотосинтезе (Благовещенский, 1940) и дыхании (Страчицкий, Рубин, 1936).
Kriedemann (1968) провел исследования фотосинтеза и дыхания листьев сорта винограда Султанина. Автор установил, что молодые, быстро растущие листья имели низкую величину чистого фотосинтеза и высокие показатели дыхания. По мере роста листьев фотосинтез устойчиво повышался (до 10 mg СO2/dm2 на час) при одновременном снижении дыхания и компенсационной точки.
Изучение интенсивности дыхания винограда в условиях Московской области для разных сортов в 1952—1953 гг. проведено Моториной (1958). Из трех сортов винограда, бывших в изучении (Альфа, Черный сладкий и Амурский), сорт Альфа отличается наибольшими величинами дыхания. В среднем интенсивность дыхания изменялась от 0,86 до 5,19 mg СO2/dm2 на час. В июне и июле дыхание составляло 50—60% фотосинтеза и снизилось в конце вегетации до 20,6—14,6%. У Амурского максимум дыхания достигал 11,1 mg СO2/dm2 на час; в конце вегетации дыхание снизилось до 1 mg. В июле дыхание у этого сорта составляло 74—80% фотосинтеза. В конце июля и в августе резко снизился фотосинтез и отношение дыхание — фотосинтез осталось высоким — 30 %. Средняя величина дыхания листьев у сорта Черный сладкий самая низкая при максимуме 5,3 mg и по отношению к синтезу составляет 30—35% в первой половине вегетации, затем в июле снижается до 21 ив сентябре —до 10,9%. Из приведенных данных видно, что дыхание листьев винограда в условиях Московской области представлено довольно значительными величинами. Расход на дыхание по отношению к фотосинтезу составил в среднем 30—40%. При этом траты на дыхание большие в начале цветения и в более южных местах произрастания. К концу вегетации траты на дыхание значительно уменьшаются и доля продуктивности фотосинтеза увеличивается. Уменьшение процесса дыхания листьев во второй половине вегетации объясняется их возрастом.
По данным автора, повышение интенсивности дыхания и траты на дыхание при высокой интенсивности фотосинтеза находятся в прямой связи с энергией роста растения. Сорт Альфа характеризуется большей силой роста, более продолжительным периодом активного фотосинтеза, повышенной интенсивностью дыхания и большими тратами на дыхание по отношению к фотосинтезу, чем сорт Черный сладкий. Сорта Альфа и Черный сладкий меньше отличаются по интенсивности дыхания, чем в отношении дыхания к продуктивности фотосинтеза. По мнению автора, для более правильной оценки значения величины дыхания следует учитывать, наряду с величиной дыхания, и отношение дыхания к продуктивности фотосинтеза. Высокое отношение дыхания к фотосинтезу у сорта Альфа не сопровождается снижением общей продуктивности, так как с возрастанием дыхания увеличивается энергия роста, что приводит к большему развитию ассимиляционной поверхности. Виноградное растение в условиях Московской области развивает высокую интенсивность дыхания, неодинаковую у разных сортов винограда и превышающую величины дыхания для других мест его произрастания.
Так, в условиях Эстонской ССР (Мийдла, 1961) на дыхание винограда расходуется в среднем 4—15% ассимилятов, образовавшихся при фотосинтезе органического вещества.
Измерение дыхания в течение суток показало, что от 0 до 12 часов дня величина дыхания практически не менялась и составляла с 23 по 29/VI 1957 г. 0,60—0,61 mg СO2/h на грамм сырого вещества и 0,40—0,41 mg/h на dm2 листа. Максимум дыхания приходился на дневные часы (12—16) и составлял 0,78 mg СO2/h на грамм сырого веса и 0,53 mg/h на dm2 листа. Минимум дыхания в течение суток наблюдался с 16 до 24 часов, а именно 0,40—0,52 mg СO2/h на dm2 листовой площади.
Зависимость дыхания от возраста листьев та же, что и в других местах произрастания винограда. Так, в условиях холодных парников в июне дыхание молодых листьев составляло 0,63 среднего возраста — 0,36 и старых 0,32 mg СO2/h.
Снижение интенсивности дыхания листьев во второй половине вегетации как общее явление для разных мест произрастания, объясняемое не только возрастом, но и изменением внешней среды у сортов и видов винограда, более приспособленных к условиям Эстонской ССР, выражено отчетливее, чем у менее приспособленных. В связи с таким выводом, наряду с другими физиологическими показателями, возможно, отмечает автор, по интенсивности дыхания судить о степени приспособленности сортов винограда к конкретным условиям выращивания. В этом отношении интересно поведение раннеспелых и позднеспелых сортов винограда. Отдельные этапы жизнедеятельности кустов ранних сортов начинаются несколько раньше, чем у более поздних. В зависимости от условий местопроизрастания эти различия в начале вегетации могут почти отсутствовать. Однако позже все процессы проходят у ранних сортов быстрее и при большей активности, чем у позднеспелых.
Для примера приводим результаты (рис. 3) исследований М а р у т я н

Рис. 4. Интенсивность дыхания (1) (mg СO2/dm2 на час) и изменения содержания аскорбиновой кислоты (2) у винограда (3). В условиях Армянской ССР до 21/VII интенсивность дыхания листьев у ранних сортов выше, чем у позднеспелых. Так, в период цветения она составляет для ранних 3—3,25 mg СO2 и для поздних сортов несколько выше —2,5 mg СO2. В дальнейшем активность дыхания листьев снижется к концу июня, и в период усиленного роста вегетативной массы и ягод интенсивность дыхания резко повышается у всех подопытных сортов.
Рис. 3. Динамика интенсивности дыхания (mg СO2) листьев винограда
Ранние сорта: 1 — Сатени белый; 2 — Араксени белый Поздние сорта; 3 — Арарати; 4 — Астамашк
Примерно, 14/VII дыхание у раннеспелых сортов резко падает и интенсивность его становится ниже, чем у позднеспелых. В период созревания интенсивность дыхания листьев ранних сортов после небольшого подъема снижается к 5/VIII до 1,6—1,7 mg СO2. В это время у позднеспелых сортов активность дыхания выше — 1,9—2,0 mg СO2. Вспышка дыхания у этих сортов достигала 2,3 mg СОа (21/VIII).
М о орите (1963) установил, что сезонные изменения содержания аскорбиновой кислоты в листьях винограда и интенсивность их дыхания (рис. 4) изменяются параллельно. Первый максимум по обоим показателям наблюдается в фазе цветения. В период роста ягод имеет место некоторое снижение, а после созревания ягод — новое повышение. Под влиянием повышенной температуры (33 °С) интенсивность дыхания сильно возрастает, при этом содержание аскорбиновой кислоты снижается.
Исследования влияния удобрений довольно широко представлены за последние годы в области виноградарства. Однако работ, затрагивающих изменения в интенсивности дыхания под влиянием удобрений, недостаточно. В этом отношении представляют определенный интерес исследования Дорохова (1966) по изучению влияния минерального питания на фотосинтез и дыхание винограда. Установлено, что влияние минерального питания непостоянно в течение вегетации и особенно для разных сочетаний питательных элементов. При внесении в почву NPK растения обладали наиболее сильным дыханием и сниженной величиной продуктивности фотосинтеза. У растений на фоне фосфорнокалийных удобрений дыхание было понижено при самой высокой продуктивности фотосинтеза.
В период роста, формирования и в начале созревания ягод наиболее высоким дыханием обладали растения на фоне РК и NK.
В период полного созревания ягод растения на всех фонах питания имели довольно высокую интенсивность дыхания. Автор заключает, что наиболее эффективно для данных почвенных условий внесение фосфорнокалийных удобрений.
Работа, проведенная в 1956 г. на сорте винограда Шасла (Плотникова, 1959), включает и наблюдения за изменениями активности дыхания листьев под влиянием микроэлементов (Мо, В) на фоне минеральных удобрений (WPK). Удобрения вносили перед цветением в почву в жидком виде.
Интенсивность дыхания с 13/VII по 9/VIII снизилась в листьях варианта NPK в период цветения и роста ягод. Подъем наступил в конце июля при подготовке ягод к началу созревания, а затем наступило снова снижение. Под. влиянием микроэлементов интенсивность дыхания на протяжении периода наблюдений была выше, чем в варианте NPK. Наибольшее усиление дыхания относится к варианту NPK+Mo+В и несколько меньше у варианта NPK+ Мо. Такое же влияние оказали микроэлементы на повышение содержания в листьях хлорофилла, однако эти изменения по фазам вегетации не совпадают с изменениями интенсивности дыхания.
Мартин с сотр. (1966) провели исследования на сорте Рислинг итальянский по изучению влияния микроэлементов на интенсивность дыхания листьев при орошении. Они установили, что увеличение дыхания через 3 дня после первого полива (период цветения) в среднем составляет 13,5%, через 6 дней— 11,7% и через 3 дня после второго полива (период созревания) — 8,4% контроля. После первого полива через 3 и 6 дней наибольшее увеличение интенсивности дыхания относится к варианту В + Мп+Мо (27,9—20,2%) и значительно меньшее увеличение имеется после второго полива (14,8%). При одинарном внесении бора усиление дыхания через 3 дня после первого полива составляло всего 9,8 %, через 6 дней — 15,3 и через 3 дня после второго полива — 8 % контроля.
Такие же небольшие прибавки получены при одинарном внесении Мn и Мо. Другие комбинации дают меньший эффект. В большинстве случаев повышение дыхания в период созревания значительно ниже, чем при поливе в период цветения и роста. Усиление активности каталазы не совпадает с изменениями интенсивности дыхания по одним и тем же комбинациям.
В исследованиях по изучению различных физиологических и биохимических показателей виноградного растения, проведенных Мехтизаде (1965),
уделено большое внимание влиянию некоторых микроэлементов на интенсивность дыхания.
Результаты исследований показали, что действие микроэлементов на интенсивность дыхания не проявляется одинаково во всех фазах развития виноградного растения.
Применение микроэлементов перед цветением и в период формирования и роста ягод при 70 % ППВ имеет значительное влияние на повышение интенсивности дыхания в листьях виноградной лозы.
Несколько более активное действие оказывают бор и медь при влажности 50% ППВ.
В фазу созревания ягод при влажности почвы 70% бор, кобальт и медь снижают активность дыхания по сравнению с контролем. При влажности почвы 50% ППВ под влиянием меди интенсивность дыхания остается на уровне контроля, а бор и кобальт несколько снижают ее по сравнению с последним. Следовательно, для некоторой активизации окислительных процессов целесообразно применение микроудобрений, приуроченное к ранним периодам развития виноградного растения.
Бор, кобальт и медь при различной влажности почвы как при внекорневом, так и при корневом питании повышают активность каталазы. Положительное действие бора, кобальта и меди проявляется более заметно при влажности почвы 50%, чем при 70% ППВ.
В настоящее время вопросы орошения виноградников приобретают все большее значение, а в связи с этим усиливается интерес к изучению водных режимов почвы и влияния их на протекание таких процессов в клетках тканей винограда, как фотосинтез и дыхание.
Изучение влияния водных режимов почвы на интенсивность фотосинтеза и дыхания сортов Каберне, Совиньон и Карабуруну проведены Стратиенко (1969) в течение вегетации прививок винограда в сосудах при влажности почвы от 20 до 80% от ППВ с интервалами в 10% (1959—1960 гг.).
Интенсивность дыхания снижается постепенно с увеличением влажности почвы, и она выше во второй половине вегетации, чем в первой. Автор отмечает, что с повышением влажности почвы от 20—30 до 50—60% отношение истинного фотосинтеза к дыханию увеличивается с 2,66 до 5,91, при дальнейшем увеличении влажности почвы величина соотношения уменьшается. В связи с полученными результатами автор заключает, что влажность почвы 50—60% от ППВ создает лучший продуктивный углеводный баланс в прививках винограда. Снижение дыхания при излишней влажности (70—80% ППВ) вызывает снижение интенсивности фотосинтеза листьев, что влечет за собой нарушение метаболических процессов в обмене веществ растения.
С ухудшением водоснабжения виноградных прививок усиливается процесс дыхания. При этом наблюдаются продуктивный фотосинтез и незначительное снижение в листьях содержания углеводов как дыхательного материала. При дальнейшем ухудшении водоснабжения (засуха) наблюдается потеря углеводов, обеднение клеток дыхательным материалом, значительное снижение процесса дыхания. Автор отмечает, что сочетание запасов влаги и содержание углеводов определяют интенсивность дыхания.
Найденов (1973) провел изучение интенсивности дыхания листьев окольцованных и неокольцованных побегов на кустах сортов винограда Жемчуг Саба, Шасла белая, Карабурну, Алиготе, Фетяска белая, Пиногри. Кольцевание проведено в период цветения. Объектами изучения были нижние и средние листья.
Под влиянием кольцевания интенсивность дыхания листьев ослабевает, но через 2—4 недели усиливается, что автор считает патологическим явлением, связанным с отрицательным воздействием избытка различных продуктов ассимиляции и диссимиляции, скапливающихся в листьях. Активность каталазы при этом ослабевает, в особенности в листьях окольцованных побегов, соцветия которых были удалены (Алиготе).
Таблица 3
Физиологические показатели работы листьев через 12—15 дней после кольцевания

При кольцевании перегруженных (Алиготе, 4 соцветия) и среднерослых побегов (Карабурну, 2 соцветия) усиливается отток ассимилятов из листьев (табл. 3), снижается накопление сухого вещества в них, а интенсивность дыхания и активность каталазы значительно повышается. Из-за недостаточного питания гроздей развитие их ослабевает и накопление сахара снижается. При этом на окольцованных побегах питание нижних гроздей ослабевает в большей мере, чем верхних (особенно Карабурну). У обоих сортов накопление сахара в ягодах больше в верхних гроздях, чем в нижних.
Автор объясняет эти нежелательные явления длительным существованием кольца на побеге как препятствия поступлению питательных веществ из других частей куста. В связи с этим автор рекомендует проводить кольцевание в виде кругового надреза коры без снятия кольца. Восстановление нормального движения питательных веществ наступает через 7—10 дней. К этому времени усиливается интенсивность дыхания и оттока, активность каталазы почти не снижается. Следует упомянуть о необходимости строгого учета возможной нагрузки побегов соцветиями в зависимости от биологических особенностей сорта и условий произрастания. Интенсивность дыхания и активность каталазы, исходя из результатов автора, могут служить показателем внесения поправок в нагрузку гроздями при использовании кольцевания побегов различных сортов винограда.
Из исследований Мехтизаде (1965) по изучению влияния гиббереллина на интенсивность дыхания листьев винограда следует, что это высокоактивное вещество повышает интенсивность дыхания листьев винограда при концентрации 0,01 %. Повышение наблюдается уже через 5 дней после опрыскивания. Параллельно автор наблюдал также усиление йодовосстанавливающей способности тканей листьев.
Изучение влияния подвоя на интенсивность дыхания и фотосинтеза листьев привоя (Плакида с сотр., 1963) показало, что под влиянием подвоя изменения фотосинтеза и дыхания довольно значительны. Интенсивность фотосинтеза на привитых кустах выше, чем на корнесобственных (Серексия). Однако для сорта Серексия из двух подвоев — Рипария Глуари Рипария х Рупестрис 3309 первый очевидно имеет большее сродство к привою, чем 3309. Интенсивность дыхания у этих компонентов очень низкая (0,61), возможно, из-за более медленного оттока ассимилятов и задержки окислительных процессов. Для сорта Рислинг, очевидно, лучшей парой будет сочетание привоя с подвоем 3309, чем 1616, так как в период определения дыхания этот процесс вообще не обнаружен. Это дает основание предположить, что в данном случае процесс дыхания вообще занижен.
Разработка комплекса физиологических и биохимических показателей для определения лучших пар для прививок с большей продуктивностью имеет большое хозяйственное значение. Диагностика этого вопроса позволила бы отказаться от длительных полевых опытов для изучения влияния подвоя на привой. В этом вопросе безусловно имеет значение взаимное влияние разных звеньев ферментных и дыхательных систем привоя и подвоя. В этом сложном вопросе свою роль должны сыграть условия среды, требуемые каждым из компонентов, и влияние подвоя, если материал для прививки берется с давно прививавшихся кустов или с корнесобственных, произрастающих в свободной от филлоксеры зоне.
Повреждение винограда хлорозом — явление частое в некоторых районах. Основной причиной, вызывающей хлороз, нужно считать нарушение обмена веществ, ведущее к полному ослаблению растения. По данным Мазанашвили (1953) при повреждении хлорозом у сильно хлорозных лоз по сравнению со здоровыми интенсивность дыхания увеличивается, возрастает активность пероксидазы в корнях и листьях, уменьшается титруемая кислотность. При этом идет интенсивная потеря пластических веществ.
Различные заболевания листьев вызывают усиление интенсивности дыхания с большой затратой энергетического материала тканей растения, быстрого его ослабления и гибели.
Durakov (1966) отмечает, что на листьях винограда в середине лета нередко проявляется хлороз, вызываемый недостатком магния. Заболевание сопровождается усилением поглощения O2 и выделением СO2. Интенсивность дыхания резко возрастает, нарушается обмен веществ, что находит отражение в изменении содержания магния, кальция, калия в пластинке и черешке листа. Определение поглощения этих веществ в черешке к содержанию их в пластинке больных листьев сравнительно с нормальными может дать указание об изменении обмена веществ при хлорозе, вызванном недостатком магния.
Весьминьш (1966) показал изменение интенсивности дыхания листьев неустойчивого к возбудителю милдью (Plasmopara viticola Ве1. et de Тoni) сорта винограда Шасла белый и устойчивого Рипария X Рупестрис 101-14. Заражение листьев возбудителем заболевания вызывало временную стимуляцию дыхания у обоих сортов. В первые дни после заражения в листьях сорта Шасла белый при температуре 25—27°С в пять раз повысилась активность полифенолоксидазы. В дальнейшем после прохождения инкубационного периода интенсивность дыхания листьев резко падала у восприимчивого сорта к мильдью Шасла белый, а у устойчивого Рипария X Рупестрис 101-4 проходил второй подъем интенсивности дыхания. В конце инкубационного периода интенсивность дыхания у сорта Рипария X Рупестрис 101-14 на протяжении 12 дней резко менялась и на листьях образовались мелкие некротические пятна, типичные для устойчивого сорта. Для сорта Шасла белый отмечена в этот период более высокая активность полифенолоксидазы в пораженных листьях по сравнению с Рипария X Рупестрис 101-14 и по сравнению с активностью пероксидазы, которая была выше у этого сорта, чем активность полифенолоксидазы. Ответная реакция на воздействие инфекции различна у неустойчивого и устойчивого сортов к мильдью и отражена в различиях прохождения этого процесса и его интенсивности.
Влияние фунгицидов на дыхательный газообмен виноградного растения почти не изучено. С этой целью Купарашвили и Чхеидзе (1971) провели полевые опыты на 6-летних кустах винограда сорта Ркацители. В исследования были включены препараты, показавшие высокую эффективность в борьбе против мильдью. В первые два дня после каждой обработки поликарбацином и цинебом интенсивность дыхания повышалась неравномерно по годам и срокам опрыскивания. На 5-, 8-й день она несколько снижалась, а в дальнейшем в течение 10—15 дней снова заметно повышалась по сравнению с контролем — для поликарбацина от 3,9 до 25% и цинеба с 6,1 до 18,7%. Интенсивность дыхания после обработки 1 %-ной бордосской жидкостью после каждого срока опрыскивания несколько активизировалась, а затем наблюдалось некоторое угнетение, после чего наступало медленное восстановление до нормы. Авторы делают вывод, что изучавшиеся два препарата можно расценивать как физиологически активные по отношению к виноградной лозе в рекомендуемых концентрациях.
В исследованиях Микеладзе (1961) у прививок Ркацители на Рипария X Рупестрис 3309 по сравнению с Ркацители корнесобственным интенсивность дыхания в большинстве случаев снижается. В привитом Саперави по сравнению с Саперави корнесобственным дыхание повышается, на основании чего Микеладзе делает вывод, что привитые лозы сильнее повреждаются хлорозом, чем корнесобственные. К неблагоприятным изменениям в прививках автор относит повышение интенсивности дыхания под влиянием подвоя (Саперави) в связи с возрастанием деятельности пероксидазы и усилением транспирации и расхода пластического материала. Следует добавить к этому, что у разных компонентов прививок изменения в обмене веществ будут проходить более или менее по-разному. Отсюда наблюдения за такими показателями, как дыхание, активность ферментных систем и др. необходимы при оценке качества прививок и выяснении взаимного влияния подвоя на привой (табл. 4).
При ослаблении жизненности организма происходит десорбция ферментов с повышением при этом их активности (Кружилин, 1955; Чрелашвили, 1951, цит. по Микеладзе, 1961).
Таблица 4
Количество поглощенного листьями O2 (cm3 на 1 g сухого вещества за 1 h)
Варианты | 16/VII 1954 г. | 4/Х 1954 г. | 22/VI 19655 г. | ||
9 ч. | 2 ч. | 9 ч. | 2 ч. | 9 ч. | |
Ркацители непривитой | 1,68 | 2,52 | 0,84 | 0,66 | 1,50 |
Ркацители на 3309 | 1,86 | 2,46 | 0,78 | 0,72 | 1,32 |
Саперави непривитой | 1,44 | 1,26 | 0,84 | 0,72 | 1,20 |
Саперави на 3309 | 1,86 | 1,62 | 0,96 | 0,75 | 1,80 |
3309 непривитой | 1,56 | 1,92 | 0,66 | 0,48 | 1,26 |
Высокой активностью пероксидазы характеризуются не только явно хлорозные лозы, но также и те из сортов подбойных американских лоз, которые считаются менее хлорозостойкими. Показатель общей активности окислительных ферментов под влиянием подвоя у Ркацители снижается, а у Саперави повышается. У корнесобственного Ркацители этот показатель значительно выше по сравнению с Саперави. Листья подвоя Р х Р 3309 по этому же показателю приближаются к Ркацители, но обладают более выраженной пероксидазной активностью (по сравнению с полифенолоксидазной).
Листьям сорта Саперави, пишет автор, по сравнению с Ркацители, свойственна также большая доля участия пероксидазы в соотношении пероксидазной и полифенолоксидазной активности (табл. 5).
Таблица 5
Активность пероксидазы и полифенолоксидазы ( cm3 KSO3 1/200 N на 1 g сухого вещества за 2 min)
