Корням принадлежит сложная физиологическая роль в процессе поглощения корневой системой воды и минеральных веществ и взаимодействия с почвенной микрофлорой. Вся деятельность корневой системы взаимосвязана с деятельностью надземных органов. В этом комплексе решающее значение принадлежит сложным биохимическим превращениям и, в частности, биологическому окислению или дыханию клеток, которое представляет собой основной источник энергии для поглотительной деятельности корней.
Дыхание клеток корней проходит при достаточной обеспеченности почвенной среды кислородом. Недостаток кислорода подавляет дыхание и все его сопровождающие системы, в результате чего снижается поглотительная активность корней. В конечном счете угнетается работа листьев, сокращается образование органических веществ, идет процесс распада их за счет внутриклеточного дыхания. Односторонность процессов обмена веществ приводит к ослаблению всего организма.
По данным Чкуасели (1964) зона тонких всасывающих корней характеризуется наибольшей интенсивностью дыхания, не падающей даже зимой и сохраняющей высокий уровень во все периоды развития. Ферментативная активность также высокая, что указывает на большую физиологическую и биохимическую активность клеток данной зоны корней.
В кончиках корней (зона меристемы) высокий уровень дыхания и активность ферментов наблюдаются в фазе роста корня. Зимой, в период покоя, когда деление клеток меристемы не наблюдается и прекращены процессы роста, интенсивность дыхания кончиков корней заметно падает. Наиболее низкими уровнем дыхания и ферментативной активностью характеризуется зона толстых проводящих корней.
Параллельно автором изучен вертикальный градиент дыхания и активности ферментов — полифенолоксидазы, аскорбиноксидазы, цитохромоксидазы, пероксидазы и каталазы для сортов подвоев — 5ББ, 420А, Рупестрис дю Ло, 3309, 101-14 и 41-Б.
О сопряженном изменении интенсивности дыхания корневой системы и побегов винограда по фенофазам развития винограда из литературных источников известно очень мало. В связи с этим исследования Мелконяна и Саркисовой (1968) сортов Воскеат и Арарати (см. раздел Побеги) несомненно заслуживают внимания.
В период сокодвижения (конец марта и начало апреля) в корнях наблюдается высокая интенсивность дыхания, которая коррелирует с интенсивностью дыхания в надземной системе (однолетние побеги). В конце апреля (период распускания почек) пониженная интенсивность дыхания корней сопровождается параллельным снижением дыхания в однолетних побегах.
Повышение интенсивности дыхания корневой системы совпадает с фазой цветения (июнь), после чего наступает постепенный спад до второй декады августа, что наблюдается и в тканях однолетних побегов. Следующее повышение интенсивности дыхания корней наступает в середине сентября и совпадает с фазой созревания ягод. В конце сентября — съемная зрелость сорта Арарати. Этот скачок дыхания автор объясняет повышением ростовых процессов в корнях, что способствует быстрой транслокации воды и продуктов метаболизма к растущим частям гроздей и ягод. В побегах продолжается постепенное снижение интенсивности дыхания по мере затухания в них ростовых процессов. После некоторого спада интенсивность дыхания корней снова повышается в период усиления созревания побегов и подготовки растения к зиме, что связано с транслокацией пластических веществ и их превращениями.
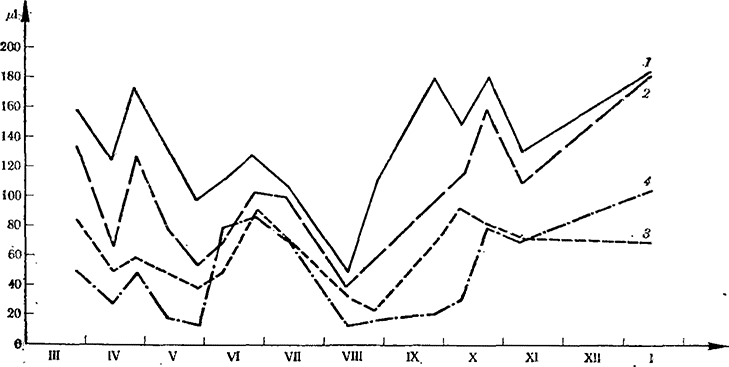
Рис. 5. Интенсивность дыхания (μl СO2/h)) по толщине корней виноградной лозы сорта Арарати в течение круг ого года
1 — всасывающие корни; 2 — мелкие корни; 3 — средние корни; 4 — толстые корни
Усиление дыхания в корневой системе происходит в осенне-зимний период (декабрь—январь). Наибольшей интенсивностью дыхания отличаются активно всасывающие корешки. По мере утолщения (рост и возраст) интенсивность дыхания корней резко ослабевает (рис. 5).
В период сокодвижения интенсивность дыхания всасывающих корней достигает 170—130, мелких 138—90, средних 88—75 и толстых — 50—70 μl. Такое снижение в зависимости от толщины корней сохраняется в течение всей вегетации. Наивысшая интенсивность дыхания отмечена для всех корней в весенний и осенний периоды. В апреле, июне, июле, августе наблюдается резкое снижение интенсивности дыхания всех корней, а у волосков и мелких корней также в октябре и ноябре. Интенсивность дыхания волосков и мелких корней достигает максимума уже в октябре — ноябре и в январе. Осенне-зимний максимум совпадает с периодом листопада, передвижением питательных веществ и осенней закалкой.
Полученные результаты иллюстрируют связь надземной части куста с его корневой системой, взаимозависимость процессов роста и метаболизма целостного растения.
В результате авторы заключают, что в течение года (март—январь) наблюдаются некоторые закономерности в процессе дыхания побегов и корней винограда. Интенсивность дыхания корней проходит волнообразно (6 волн повышения) и коррелирует с основными фазами развития и органогенеза надземных органов куста. Дыхание корней находится в тесной зависимости от их возраста. По величине интенсивности дыхания выделяются всасывающие корни, интенсивность дыхания у которых снижается по мере их утолщения. Проведенные исследования показывают взаимосвязь по интенсивности дыхания между корневой системой и надземной частью куста, что подчеркивает существующую связь этих двух частей растения в общем метаболизме веществ.
Изучение влияния состава питательной среды на дыхание корней виноградной лозы провел и Киквидзе (1966). Автором исследовалось дыхание корней у двух подвойных сортов виноградной лозы — Берландиери х Рипария Кобера 5ББ и Рипария х Рупестрис 101-14, помещенных в раствор Кноппа с выключением, либо изменением содержания отдельных солей. Дыхание корней у 101-14 во всех случаях, предусмотренных опытом, было более интенсивным, чем у 5ББ. Выключение отдельных элементов из питательной среды вызвало снижение дыхания корней у обоих сортов. Наиболее резко падало дыхание, когда из питательного раствора был исключен фосфор. Дыхание значительно падало также при исключении солей калия и магния. С другой стороны, повышение концентрации питательного раствора (особенно повышение концентрации калия) также вызывало снижение дыхания корней. Однако увеличение содержания фосфора оказывало иное влияние: у корней 5ББ снижение дыхания не было столь значительно, а у корней 101-14 оно даже возрастало, притом очень интенсивно.
Рост и развитие корней, как и каллюса, сопровождается окислительными процессами, среди которых значительное место занимает дыхание. Процесс дыхания неразрывно связан с затратами пластических веществ на новообразования молодых тканей каллюса и корешков. При стратификации для дыхания основное значение принадлежит температуре. В связи с этим необходимо знать, как влияет изменение температуры в стратификационных ящиках на процесс дыхания тканей корешков и каллюса.
В 1959 г. было проведено изучение влияния различной температуры на интенсивность дыхания тканей корешков и каллюса прививок винограда и сопоставление полученных данных с расходом ими пластических веществ (Мишуренко и др., 1961). Через 18 дней после помещения прививок на стратификацию в отобранных пробах определяли интенсивность дыхания на приборе Варбурга при 15, 16, 25 и 30 °С (экспозиция 60 min, навеска сырого материала 500 mg). Оказалось, что при одной и той же температуре (25 °С) интенсивность дыхания неодинакова у различных сортов винограда.
У каллюса привоя и подвоя, если в качестве привоя были взяты черенки сортов Королева виноградников и Алиготе, интенсивность дыхания была в полтора раза выше, чем в том случае, когда привоем служил Каберне Совиньон. При температуре 15—16°С такой разницы не наблюдалось.
У корешков подвоя при использовании в качестве привоя черенков сорта Каберне Совиньон дыхание было несколько интенсивнее в сравнении с черенками сортов Королева виноградников и Алиготе.
Различия в интенсивности дыхания каллюса и корешков разных сортов можно объяснить их биологическими особенностями, а также, очевидно, аффинитетом к взятому подвою. С понижением температуры интенсивность дыхания снижается, при температуре 9—10°С дыхания не обнаружено.
Снижение интенсивности дыхания при понижении температуры обосновывает физиологически снижение затрат пластических веществ, а также целесообразность температурного режима при стратификации у основания подвоя.
Интенсивность дыхания корешков подвоя при 25 и 30 °С в 1,5 раза выше, чем у корешков привоя. Интенсивность дыхания каллюса подвоя при 25 °С в большинстве случаев ниже, чем у каллюса привоя, а при 30°С, наоборот, выше у каллюса подвоя. Интенсивность дыхания каллюса пятки подвоя, примерно, такая же, как и корешков подвоя, а каллюса у места спайки выше, чем тканей корешков привоя.
Из данных более ранних исследований видно, что наибольший расход питательных веществ был у прививок, стратифицированных при повышенной температуре у основания подвоя в результате усиления процесса дыхания.
Результаты проведенных исследований показали необходимость задержки развития корней и каллюса у основания подвоя при стратификации и закалке виноградных прививок, так как при высадке прививок в школку большая часть нежных корней, развившихся при стратификации, повреждается, а развитие новых корней в школке от пятки подвоя задерживается из-за недостатка пластических веществ в подвое.
Между интенсивностью дыхания, интенсивностью окислительных процессов и устойчивостью растений к заболеванию существует положительная зависимость. В ряде случаев конечный результат этого взаимодействия определяется способностью растений сохранять нормальный ход дыхательного газообмена (Рубин, 1951, 1953). При заболевании значительно активируется дыхание, в дальнейшем оно падает. Биохимические различия между здоровыми и пораженными филлоксерой корнями дают возможность судить о сдвигах в обмене веществ, которые вызывает паразит.
В связи с некоторыми особенностями дыхания и окислительных ферментов у корней винограда в отношении филлоксероустойчивости проведены наблюдения Субботиной (1961) на сортах Шасла белая (неустойчивый), Каберне Совиньон (несколько устойчивее, чем Шасла) и подвоя Рипария X Рупестрис 101-14 (РхР 101-14).
Наибольшей интенсивностью дыхания обладают корни РхР 101-14, наименьшей — Шасла белой, а Каберне Совиньон занимает промежуточное место. К концу вегетации (созревание ягод) снижается дыхание у устойчивых сортов РхР 101-14 и Каберне. У Шасла интенсивность дыхания в этот период довольно высокая и выше, чем в период цветения (табл. 8).
Возрастание интенсивности дыхания и окислительных ферментов является признаком, характерным для заболевания.
Шасла белая и РхР 101-14 резко отличаются по активности каталазы. У Шасла в течение вегетации активность каталазы значительно выше, чем у РхР 101-14. У Каберне в начале вегетации она занимает промежуточное место, а к концу вегетации сильно падает.
Таблица 8
Активность дыхания корней винограда (cm СO2/g сырого веса на час)
Сорт | Корни | Перед | Цветение | Созревание |
Шасла белая | крупные | 90,5 | 66,2 | 73,4 |
| средние | 78,8 | 30,2 | 73,2 |
| мелкие | 144,9 | 23,6 | 96,4 |
Каберне Совиньон | крупные | 132,2 | 54,5 | 27,6 |
| средние | 127,1 | 91,0 | 31,3 |
| мелкие | 73,3 | 66,1 | 18,2 |
Рипария х Рупестрис 101-14 | крупные | 229,3 | 35,2 | 44,5 |
| средние | 278,7 | 236,5 | 55,3 |
| мелкие | 145,1 | 120,4 | 60,1 |
Наибольшая активность полифенолоксидазы установлена у Каберне, наименьшая — у РхР 101-14. Корни Шасла обладают вдвое большей активностью в течение вегетации, чем РхР 101-14. Как видно, отмечает автор, характер изменений обмена связан со степенью устойчивости сорта. Поражение корней винограда филлоксерой вызывает повышение интенсивности их дыхания. Наиболее велика интенсивность дыхания РхР 101-14. У сорта Шасла белая в течение вегетации наблюдается повышение активности окислительных ферментов (каталазы и полифенолоксидазы). У подвоя РхР 101-14 активность окислительных ферментов значительно ниже, чем у Шасла.
Согласно теории Рубина и Арциховской (1960) повышение общего уровня энергетического обмена в пораженных тканях и связанное с этим усиление синтеза обусловливается перестройкой дыхательного аппарата путем замены нестойких систем на стойкие, способные осуществлять свои функции в условиях неблагоприятного воздействия паразита. Такая теория требует исследований в направлении раскрытия природы биохимического процесса, обусловливающей филлоксероустойчивость. В этом направлении интересны работы Зотова (1964), выводы которых рассматриваются в разделе по устойчивости.
В последние годы, отмечают Джапаридзе и Чкуасели (1972), советскими учеными показана прямая зависимость между потребностью в кислородном питании корней и проявлением эдафизического хлороза. В связи с этим получает большое значение знание сортовых различий корней в потребности кислорода, на что уже ранее указывалось. В связи с этим авторами была изучена интенсивность дыхания двух подвойных гибридов виноградной лозы: хлорозостойкого Берландиери х Рипария Кобер 5ББ и нестойкого Рипариях Рупестрис 3309 (РхР 3309).
Динамика дыхания лозы в течение вегетационного периода показала, что корни гибрида РхР 3309 дышат в 1,5—2 раза интенсивнее, чем корни гибрида Б х Р Кобер 5ББ. В период зимования анализы показали значительное снижение уровня дыхания, однако сортовые различия при этом сохраняются.
При снижении содержания кислорода в почве сильнее страдают от гипоксии корни более требовательного сорта РхР 3309, так как у них нарушается согласованность различных функций, что легко переходит в состояние хлороза.
Гибрид Берландиери х Рипария Кобер 5ББ (Б х Р Кобер 5ББ) расходует относительно меньшее количество свободного почвенного кислорода, что сочетается с его более высокой хлорозоустойчивостью. К этому следует добавить, что, очевидно, у гибрида БхР Кобер 5ББ комплекс окислительновосстановительных ферментов более активный, вырабатывающий более специфическую ответную реакцию, препятствующую появлению хлороза.