Брожение в присутствии уксусной кислоты
При рассмотрении процентного содержания уксусной кислоты, образовавшейся во время спиртового брожения, приходится констатировать, что она образуется неравномерно сброженному сахару. Вначале ее образование идет быстро, потом замедляется; к концу брожения содержание кислоты может даже уменьшиться, иногда сильно (Вантер, 1937; Пейно, 1939 и 1946). Это явление протекает по-разному в зависимости от вида дрожжей (см. рис. 15.17). Более того, дрожжи способны поглощать уксусную кислоту, которую добавляют перед брожением (рис. 15.18).
Можно допустить, что такое использование уксусной кислоты дрожжами включает, прежде всего, ее восстановление в этаналь, которое способствует спиртовому брожению в ущерб брожению глицерино-пировиноградному, увеличивая образование спирта, а также вторичных продуктов при одновременном уменьшении содержания глицерина. На деле констатируют, что добавление уксусной кислоты снижает уровень глицерина и повышает содержание 1бутандиола-2,3 и ацетилметил- карбинола.

Рис. 15.18. Уменьшение содержания уксусной кислоты, добавляемой в сусло до брожения:
1 — дрожжи М; 2— дрожжи О; 3 — дрожжи С; 4—дрожжи S; 5 — дрожжи В.
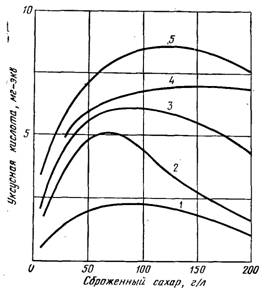
Рис. 15.17. Образование уксусной кислоты в зависимости от сахара, сбраживаемого пятью различными видами дрожжей:
1 — контроль; 2 6,2 мг-экв уксусной кислоты; 3 h 12,4 мг-экв уксусной кислоты.
Косвенным следствием этого свойства дрожжей разлагать уксусную кислоту является возможность снижения путем повторного брожения содержания летучей кислотности в больном вине. Если к виноградному суслу или к собранному винограду добавлять вино, обладающее повышенной летучей кислотностью, разумеется, после стабилизации его с точки зрения микробиальной активности в соотношении от 1/3 до 1/2 так, чтобы летучая :кислотность смеси была порядка 0,60 r/л, то после брожения получают вино, в котором содержание уксусной кислоты обычно не (Превосходит уровня, соответствующего нормальному брожению. Одновременно снижается процентное содержание этилацетата. Понятно, что такой способ может быть разрешен только для вина, летучая кислотность которого не превосходит установленного предела (0,90г/л).
Разложение яблочной кислоты
Давно известно, что содержание яблочной кислоты в виноградном сусле во время спиртового брожения уменьшается независимо от яблочно-молочного брожения. Пейно (1938) показал, что это общее явление и что в зависимости от вида дрожжей потери яблочной кислоты составляют от 10 до 24% ее первоначального количества; разложение тем более значительно, чем ниже рН, и при рН 5,0 оно прекращается. Но содержание яблочной кислоты в (вине, хранившемся в течение года в 'присутствии дрожжей, .не изменяется; уменьшение содержания этой кислоты происходит исключительно в период брожения.
Пейно и сотрудники (,1964) и Пейно и Сюдро (1964) показали, что некоторые дрожжи рода Schizosaccharomyces обладают свойством разлагать в анаэробиозе значительные количества яблочной кислоты (до 90%) (Schizosaccharomyces pombe).
Каков бы ни был тип рассматриваемых дрожжей, разложение яблочной кислоты соответствует по своему характеру истинному брожению согласно общему уравнению.
СООН — СНОН - СН2 - СООН - I _> сн3 — СН2ОН + 2С02.
Фук и Радлер (1972) показали, что в первое время яблочная кислота превращается в пировиноградную кислоту (ом. рис. 15.27), пировиноградная кислота затем участвует в реакциях спиртового брожения (см. рис. 15.12) с декарбоксилированием в этаналь, который сам .восстанавливается в этанол.
Метаболизм азотистых компонентов и образование высших спиртов
Проблемы питания дрожжей азотом и превращений азотсодержащих компонентов винограда во время брожения хорошо известны (Дюпюи и сотрудники, 1967) и описаны в гл. 7.
Синтез аминокислот и белков дрожжами основан на классических механизмах биохимии. Аммиачный азот поступает из а-кетоглутаровой кислоты, чтобы дать глутаминовую кислоту (рис. 15.19). Все другие аминокислоты образуются или изменением карбоновой структуры глутаминовой кислоты, например при образовании пролина (рис. 15.20), или же трансаминированием, например путем синтеза аспарагнновой кислоты из щавелево-уксусной или синтезом аланина из пировиноградной кислоты (рис. 15.21).
Среди продуктов метаболического разложения аминокислот наибольшее значение для энологии, безусловно, имеют высшие спирты: действительно, они участвуют как сами по себе, так и в виде своих сложных эфиров в образовании органолептических характеристик вин.
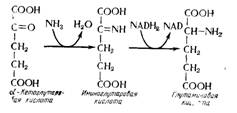
Рис. 15.19. Образование глутаминовой кислоты из минерального азота.
С момента опубликования фундаментальных работ Эрлиха известно, что аминокислоты являются главными предшественниками высших спиртов. Механизм этой реакции, который специально исследовали Сенте Шанмуганатан и Эльсден (1958), включает дезаминирование аминокислоты в кетокислоту, которая декарбоксилируется в альдегид, в свою очередь восстанавливающийся в спирт (рис. 15.22).
В табл. 15.3 показано соответствие между аминокислотами с л+1 атомами углерода и спиртами с п атомами углерода. Роль кетоновых кислот, или, точнее, альдегидов, которые являются их производными, .как акцепторов водорода :в форме NADH2, получающийся при гликолизе, по-видимому, указывает на то, что высшие ширты должны участвовать вместе с глицерином в составлении баланса продуктов глицерино-пировиноградного брожения (раздел «Баланс вторичных продуктов глицерино-пировиноградного брожения») .
Но, как заметили Дюпюи и сотрудники (1967), в бродящих средах содержатся высшие спирты, например пропанол-1, которые не имеют возможных предшественников среди натуральных аминокислот (см. табл. 15.3). Кроме того, не установлено зависимости между содержанием аминокислот и содержанием соответствующих высших шортов. Помимо этого не существует связи между образованием высших спиртов и азотистым питанием дрожжей, например когда проводят несколько последовательных брожений одними и теми же не размножающимися делением дрожжами без внесения азотистого питания, то констатируют, что образование высших спиртов каждый раз происходит в количестве, пропорциональном содержанию сахара. Эти наблюдения подтверждают гипотезу, выдвинутую Пейно и Гимберто (1958 и 1959), согласно которой реакция Эрлиха объясняет образование лишь части высших спиртов.

Рис. 15.20. Образование пролина за счет изменения молекулы глутаминовой кислоты.

Рис. 15.21. Образование аспарагиновой кислоты и аланина из глутаминовой кислоты путем трансаминирования.
По этому же вопросу важные факторы сообщили Ингрээм и Гимон (1960), Ингрээм и сотрудники (1961) и Гимон (1966). Они провели серию оригинальных экспериментов, используя [мутанты Saccharomyces cerevisiae, потерявшие способность сбраживать некоторые аминокислоты. Авторы констатировали, что Эти самые штаммы также потеряли способность синтезировать соответствующие высшие спирты и подтвердили существование взаимосвязи между синтезом этих спиртов и синтезом аминокислот. Но в то же время синтез других высших спиртов, в частности пропанола-1, бутанола- и гексанола-1, для которых не су-, ществует аминокислот-предшественников, возрастает по отношению к первоначальному штамму. От этого штамма получен мутант. Таким образом, типы синтеза различных высших спиртов связаны общими механизмами, в которых участие аминокислот необязательно.
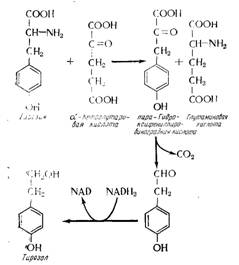
Рис. 15.22. Механизм биосинтеза высших спиртов из соответствующих аминокислот: образование тирозола из тирозина.
Из своих работ авторы заключают, что обязательными предшественниками высших спиртов являются не аминокислоты, а соответствующие кетоновые кислоты; именно их синтез ингибируется у мутантов. Эти кетоновые кислоты получаются не только от разложения аминокислот в соответствии с реакцией Эрлиха (ом. рис. 15.22), но и в результате углеводного метаболизма. Отсюда следует, что некоторые высшие спирты (изоамиловый, изобутиловый) могут иметь несколько механизмов синтеза (из аминокислот или путем углеводного метаболизма), тоща как другие образуются только одним способом (углеводного метаболизма). Взаимообратимость этих двух путей обмена веществ описана, в частности, в публикации Гимона (1966). В качестве примера можно указать на механизм образования я-пропилового спирта (рис. 15.23), для которого не существует соответствующей аминокислоты.
В этом случае уксусная кислота участвует в виде ацетилкофермента А. Так же, как можно перейти от пировиноградной кислоты с Сз к а-кетобутановой кислоте с С4, можно представить переход от последней к кетоновым кислотам с С5, Сб или С7 и написать образование я-бутиловых, «- амиловых (пентанол-1) и к-гексиловых спиртов. Этот механизм применим к разветвленным кетоновым кислотам и позволяет объяснить возможность синтеза основных высших спиртов (изобутилового, изоамилового и амилового активного) без вмешательства соответствующих аминокислот. Кроме того, гексанол вина должен происходить из альдегидов с ;Се (гексаналь, гексенал), присутствующих в винограде, как и во всех растениях.
Образование высших спиртов и их сложных эфиров зависит от штамма дрожжей, условий среды и аэрации (Бертран и Риберо-Гайон, 1968).