Морфологические, анатомические и физиологические особенности стебля винограда
Стебель винограда - это осевой надземный вегетативный орган, который обеспечивает двухстороннее передвижение воды и питательных веществ по растению. Стебель поддерживает листья, располагая их в условиях максимальной освещенности, и участвует в накоплении воды и запасных питательных веществ в растении. Стебель представляет собой ось виноградного побега, причем у молодых побегов он травянистый и несет на себе листья, пасынки, усики, соцветия и другие боковые органы. С возрастом он одревесневает, образуя скелет куста в виде многолетних надземных частей (штамб, рукава, рожки, плодовые звенья и др.). В отличие от других древесных и кустарниковых растений стебель винограда не имеет осевой симметрии и постоянной формы. Длина и толщина стебля винограда может варьировать в значительных пределах. У дикого винограда стебель имеет вид, длинного (8-10 и до 30 м), гибкого оголенного ствола, взбирающегося вверх по деревьям. На ровных местах он свободно стелится по земле.
В культуре виноградное растение приобретает форму куста со стеблем разных очертаний (прямостоячий, наклонный, с головкой, рукавами и др.). Толщина стебля может достигать 70-80 см и более. Так, в Калифорнии зарегистрирован 175-летний куст с окружностью ствола 275 см.
Стебель однолетнего возраста состоит из узлов и междоузлий. На узлах располагаются листья, пасынки, зимующие глазки, соцветия и усики (рис. 8, А). Соцветия затем превращаются в грозди, а с помощью усиков стебель прикрепляется к опорам. Листья размещаются на узлах супротивно-переменно по формуле 1/2. Однако у сеянцев листорасположение от основания до первого усика выражается формулой 2/5. Внутри каждого узла имеется перегородка-диафрагма (рис.8, Б). На междоузлиях побега никакие органы не развиваются. В пазухах листьев образуются почки, из которых в течение того же вегетационного периода развиваются побеги второго порядка - пасынки. В пазухе самого нижнего пасынкового листа формируется почка, отличающаяся крупными размерами - это зимующий глазок. Из зимующих глазков образуются основные побеги лишь весной следующего года. Выше 3-5 узла, против листьев, образуются усики. У плодоносных побегов вместо усиков, на самых нижних узлах (от 2 до 6-го), формируются соцветия. Количество соцветий на одном побеге варьирует в зависимости от сорта, условий произрастания и питания, от 1-3 до 6-7.
Усики располагаются в определенном порядке: 2 узла с усиком, один без усика. Исключение составляет вид V. labrusca (рис.8, Д, 2). Прерывистое расположение усиков и соцветий по длине побега винограда объясняется характерной сменой их моноподиального и симподиального роста (рис. 8, Г, Д). Первоначально между 2-5 узлами главная ось побега растет только верхушечной точкой роста. Это, так называемый, моноподиальный рост. Начиная с 3-6 узла моноподиальный рост сменяется симподиальным, когда основная ось побега смещается и уступает место новому бугорку, который продолжает рост до следующего узла, где опять новая точка роста смещается и образуется новый бугорок, продолжающий рост побега. Смещенные точки роста образуют соцветия или усики. Следовательно, соцветие или усик - это редуцированные верхушки побега, смещенного в сторону.
Развитие стебля может происходить и из зародышевой почки семени. В этих случаях моноподиальный рост стебля продолжается до 6-10 узла, а затем начинается чередование моноподиального роста с симподиальным.
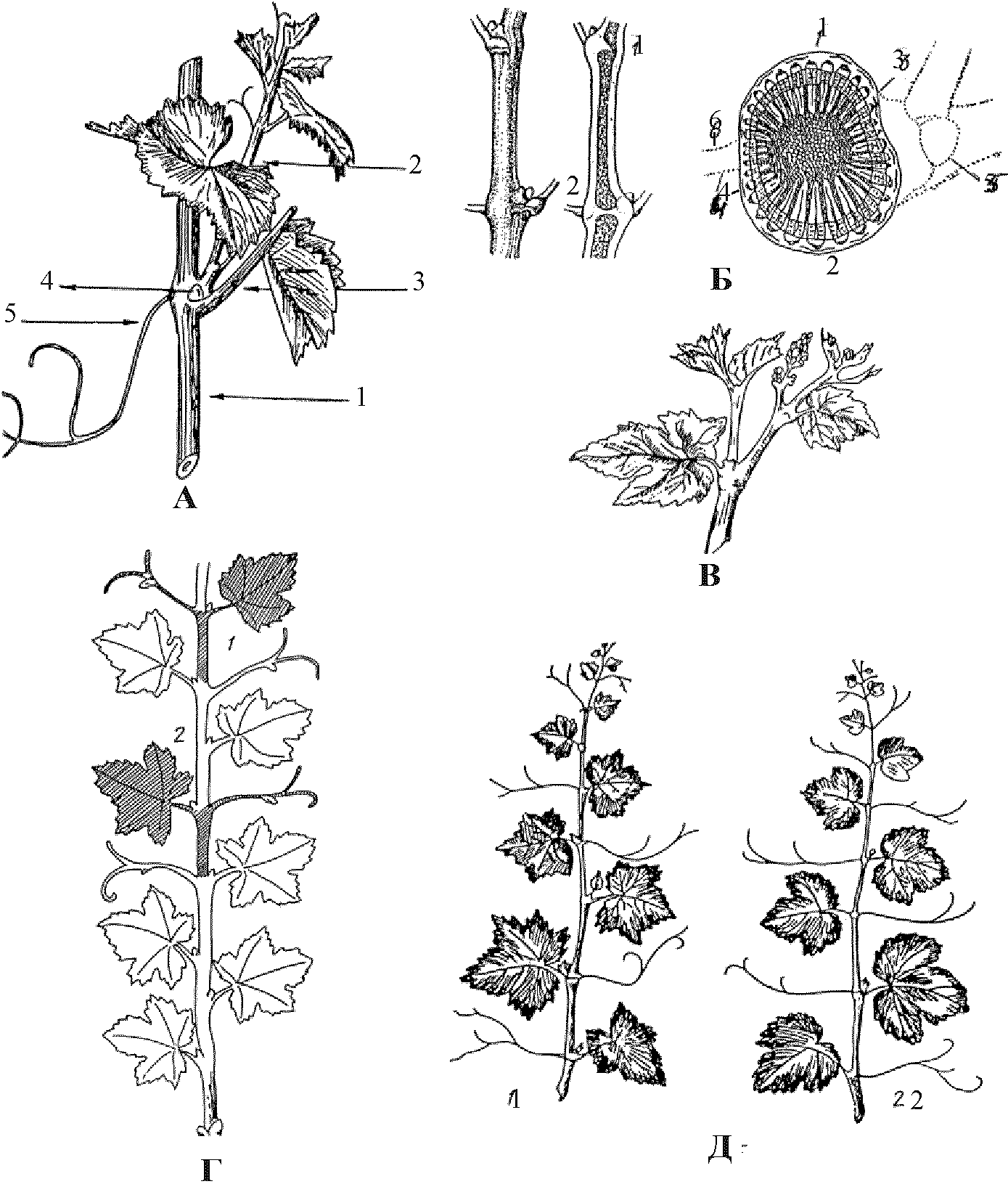
Рис. 8. Морфология побега
А — морфология узла однолетнего побега (1 - основной побег; 2 - пасынок или побег второго порядка; 3 - черешок листа; 4 - зимующая почка; 5 - усик).
Б - продольный (слева) и поперечный (справа) разрез однолетнего побега винограда (1 - диафрагма на узле без усика - прерывистая; 2 - диафрагма на узле с усиком - сплошная; 1 - спинная сторона побега; 2 - брюшная сторона побега; 3 - желобчатая сторона побега; 4 - плоская сторона побега; 5 - зимующий глазок; 6 - усик ).
В — внепазушный побег.
Г — схема побега винограда V. Vinifera L., показывающая симподиальное (1) и моноподиальное (2) ветвление.
Д — характер расположения листьев и усиков (1 - прерывистое; 2 - непрерывное).
Однолетний побег винограда имеет ассимметричное (дорзивентральное) строение, то есть имеет спинную, брюшную, желобковую и плоскую стороны (рис. 8, Б), которые отличаются степенью развития, интенсивностью физиолого-биохимических процессов. К спинной стороне смещены все боковые побеги (пасынки), а к брюшной - все почки (зимующие глазки). Брюшная сторона слабее окрашена и всегда сильнее развита, чем остальные. Желобчатая сторона начинается от зимующего глазка и тянется вверх до следующего узла. Сторона побега, расположенная против желобчатой, называется плоской. На этой стороне желобок значительно менее выражен, чем на желобчатой. Желобчатая и плоская стороны непрерывно чередуются, а спинная и брюшная стороны по всей длине побега сохраняют одинаковое пространственное положение. Так как на спинной стороне рост клеток в продольном направлении происходит сильнее, чем на брюшной, то верхушка побега изгибается. Чем сильнее рост, тем больше изгиб. Выпрямление верхушки указывает на прекращение роста стебля в длину. Изгиб верхушки побега является приспособлением для защиты ее от механических повреждений.
Верхушка побегов со стоит из меристематических тканей, имеет полусферическую форму и защищена развивающимися зачаточными листочками, плотно ее прикрывающими.
Анатомическое строение стебля винограда тесно связано с выполняемыми им функциями. Для него характерно как первичное, так и вторичное строение.
Первичное анатомическое строение является результатом дифференциации клеток первичной меристемы в конусе нарастания на верхушке побега. На поперечном срезе стебля молодого побега можно увидеть под микроскопом эпидермис, первичную кору и центральный цилиндр (рис. 9, А).
- Эпидермис - это защитная ткань, состоящая из многоугольных плотно расположенных клеток, содержащих значительное количество танина и имеющих многослойную оболочку покрытую кутикулой. В зависимости от сорта и условий произрастания оболочка может иметь различные выросты (волоски, жемчужные железки).
- Первичная кора включает 8-10 слоев коровой паренхимы из округлых плотно расположенных клеток, содержащих крахмал, сахар, танин, хлоропласты, один слой эндодермы (самый внутренний слой). В первичной коре образуется колленхима - механическая ткань, состоящая из клеток имеющих более толстые стенки. Колленхима располагается напротив сосудисто-волокнистых пучков центрального цилиндра.
- Центральный цилиндр состоит из перицикла (снаружи), проводящих пучков (чаще их 5), расположенных радиально, и развитой сердцевины. Между пучками образуются первичные сердцевинные лучи . Над пучками находятся склеренхимные (перекамбиальные, перецикловые) тяжи.
Сосудисто-волокнистые пучки содержат у периферии первичный луб, состоящий из ситовидных трубок и лубяной паренхимы, а внутри
- первичную древесину, состоящую из сосудов древесины и древесинной паренхимы. Между лубом и древесиной дифференцируются клетки камбия, который проходит и через сердцевинные лучи.
Первичная структура побега быстро переходит во вторичную (рис. 9, Б, В, Г). Как и у корня, вторичные образования зависят от деятельности двух различных образовательных тканей - камбия и феллогена.
В период вегетации камбий образует к наружи вторичный луб , а во внутрь
- вторичную древесину. Вторичная флоэма состоит из чередующихся слоев клеток мягкого и твердого луба, а ксилема - из крупных сосудов, древесинной паренхимы и древесинных волокон. Камбий, находящийся в сердцевинных лучах, откладывает и кнаружи и вовнутрь клетки сердцевинных лучей. Сердцевинные лучи возникают дополнительно и внутри пучка, которые своими концами не доходят до сердцевины. Это, так называемые, радиальные лучи.
Осенью камбий приостанавливает свою деятельность с тем, чтобы следующей весной снова начать образование новых тканей, как и в первый год. Побег утолщается, образуя годичные кольца, толщина которых зависит от условий роста стебля в течение года (0,6-3,4 мм). По кольцам можно определить возраст куста или надземной части.
В августе, в мягком лубе или в перицикле, появляется новая образовательная ткань - феллоген, дающая начало новым тканям - пробке (феллема) и феллодерме, которые вместе образуют перидерму , в связи с чем зеленая окраска побега переходит в коричневую, что является признаком его вызревания. Перидерма кроме феллогена включает еще 4-5 слоев пробки и 2-3 слоя феллодермы. В результате образования перидермы теряется связь между тканями расположенными кнаружи от нее и находящимися в середине побега, вследствие чего внешние ткани отмирают, а кора приобретает характерную окраску для вызревшего побега.
Впоследствии, ежегодно, в более глубоких зонах луба, образуются новые перидермы, которые также вызывают отмирание новых тканей, образуя на поверхности ствола так называемую корку. У некоторых сортов бывает двухслойная перидерма. Ученые связывают это с морозоустойчивостью сортов.

Рис. 9. Анатомия побега:
А — первичное анатомическое строение стебля (1 - эпидермис; 2 - колленхима; 3 - коровая паренхима; 4 - эндодерма; 5 - склеренхима или перикамбиальные тяжи; 6 - первичная флоэма; 7 - камбий; 8 - первичная ксилема; 9 - сердцевина; 10 - сердцевинный луч);
Б, В, Г — вторичное анатомическое строение стебля (1 - сердцевина; 2 - сердцевинные лучи; 3 - ксилема; 4, 5 - радиальные лучи; 6 - камбий; 7-10 - вторичная флоэма; 7 и 9 - мягкий луб; 8 и 10 - твердый луб; 11 - первичная флоэма; 12-14 - перидерма; 12 - феллодерма; 13 - феллоген; 14 - филлема; 15 - перицикл; 16 - перицикловые тяжи; 17 - эндодерма; 18 - коровая паренхима; 19 - колленхима; 20 - эпидермис).
Вторичное строение характеризуется еще и тем, что к концу лета несколько слоев мелких клеток сердцевины, расположенных непосредственно около древесинного цилиндра, в результате отложения на их стенках, пропитанных суберином пластинок, опробковевают и сердцевина, в связи с изоляцией, отмирает и буреет. Внутренний пробковый слой служит для защиты центрального цилиндра со стороны сердцевины.
Анатомическое строение узлов несколько отличается от строения междоузлий. На узлах наблюдается более мощное развитие паренхимных и механических тканей и слабое развитие сосудистой системы. Коровая паренхима, сердцевинные лучи и перицикловые тяжи на узлах шире.
К физиологическим свойствам стебля винограда относятся: рост; поглощение и перемещение питательных веществ; уровень содержания и динамика накопления пластических и физиологических активных веществ; транспирация и ассимиляция побегов и др. Изучение физиологических свойств стебля имеет большое значение для научного обоснования агротехники винограда.
- Рост стебля происходит в следующей последовательности: в верхушечной меристеме конуса роста стебля происходит деление клеток; за фазой деления наступает фаза растяжения клеток, во время которой рост происходит очень энергично. При сильном росте побегов наиболее быстро удлиняются второе и третье междоузлия, считая от верхушки, иными словами осуществляется, так называемый, интеркалярный рост. Ниже по побегу, этот процесс замедляется и на шестом-седьмом междоузлии почти полностью прекращается.
В росте побегов в течение вегетационного периода наблюдается так называемый “большой период роста”. Исследованиями многих авторов установлено, что после распускания почек молодые побеги растут медленно, потом их рост ускоряется и достигает максимума во время цветения; затем он замедляется, а после начала созревания ягод полностью прекращается. Однако в зависимости от сорта, метеорологических условий и применяемой агротехники характер “кривой роста” побегов может значительно изменяться.
Установлено, что срок наступления цветения и период максимального роста побегов меняются и по годам. Совпадение периода цветения с максимальным ростом побегов можно ожидать только тогда, когда оптимальная для роста побегов температура наступает до цветения или к началу периода цветения.
Исследования А. С. Мержаниана (1948) показали, что рост междоузлий неодинаков. У междоузлий, заключенных между узлами с усиками (симподиальное развитие), он проходит интенсивнее, чем у соседних междоузлий, имеющих моноподиальный тип ветвления.
Остановка роста происходит раньше всего в нижней, затем в средней и позже всего - в верхней части побега. Количество междоузлий, на которых наблюдается рост, может быть разным, в зависимости от условий произрастания. Весной, в начале вегетации, растет большое число междоузлий - до восьми, в то время как летом - всего лишь четыре-пять.
Помимо роста в длину, побег растет и в толщину. Первоначально это происходит за счет деления клеток, а позже - за счет растягивания их в тангентальном и радиальном направлениях. Ниже зоны наибольшего удлинения, побег утолщается за счет деятельности камбия.
Пасынки, так же как и основной побег, имеют большой период роста, который выражен неодинаково в зависимости от положения пасынка по длине побега. Рост пасынков у самого нижнего узла главного побега небольшой. У 3-5-го узлов он достигает максимума, и поэтому здесь пасынки имеют наибольшие размеры. На узлах верхней зоны рост пасынков снова уменьшается.
На рост побегов большое влияние оказывают сортовые особенности, полярность, сосущая сила, физиологическая влажность, корневое давление, различные экологические условия (температура, влажность), питание и др.
- Разные сорта обладают неодинаковой силой роста. Так, среднеазиатские столовые сорта восточной ветви (convar orientalis) характеризуются более сильным ростом, чем сорта западной ветви (convar occidentalis).
- На рост побегов большое влияние оказывает явление полярности.
При вертикальном положении побега, прежде всего, трогаются в рост глазки верхушечной зоны, позже распускаются глазки средней и нижней зон. Удаление верхушечной точки роста стимулирует рост пасынковых побегов. Рост побегов значительно сильнее при вертикальном положении, однако, ветвление при этом слабое; при горизонтальном положении рост слабый, но ветвление сильное и рост пасынков более интенсивный. При горизонтальном расположении одно-, двух- или многолетних ветвей, полярность проявляется очень слабо, и образующиеся побеги растут равномерно.
- Сосущая сила тканей побега, от которой зависит растяжение клеток, является важным фактором роста. На нее оказывают влияние транспирация и корневое давление. Известно, что чем выше корневое давление, и, чем больше жидкости вытекает при “плаче” из срезов на кусте, тем сильнее растет куст.
- Большое значение для роста имеет также повышенная физиологическая влажность побегов, что способствует улучшению снабжения всех тканей водой, необходимой для осуществления растяжения клеток.
Физиологическая влажность тканей и клеток зависит от внешних условий роста, от силы всасывания воды корневой системой и развития поглощающих корешков, а также от интенсивности транспирации, от величины и состояния проводящей системы.
- Значительное влияние на рост виноградного растения оказывают факторы внешней среды. Из метеорологических факторов доминирующее влияние на рост побегов оказывает температура. Наиболее интенсивно происходит рост растений винограда при 28- 30°С. При температуре ниже 8° виноградное растение не растет, а при температуре выше 40-42°С рост также прекращается.
Значение температурного фактора для роста побегов выявляется при сопоставлении сезонных изменений прироста и температуры. Исследованиями М.М. Никова (1962) установлено, что суточный прирост побегов идет почти параллельно величине суточной температуры, что указывает на их функциональную зависимость. Однако эта взаимосвязь устанавливается лишь в период интенсивного роста. С ослаблением темпа роста, реакция на повышение температурного фактора непрерывно уменьшается. При достаточном увлажнении почвы и одновременном наличии других факторов жизни, виноградное растение характеризуется интенсивным ростом, который может усиливаться при повышенной влажности воздуха.
- Рост винограда в значительной мере обусловлен условиями минерального питания . Установлено, что азот, фосфор и калий обеспечивают сильный рост кустов винограда. Самое сильное действие на рост виноградного растения оказывает азот, но если при этом в достаточном количестве имеются все другие элементы питания. Большое участие в процессах роста принимают и микроэлементы: бор, цинк, марганец, молибден и др.
- Рост побегов увеличивается под влиянием стимуляторов роста ( - индолилуксусная кислота, индолилмасляная кислота, - нафтилуксусная кислота и др.), а также веществ высокой физиологической активности (гиббереллин и др.). В то же время хлорхолинхлорид (ССС) подавляет рост побегов и увеличивает пасынкообразование.
- Важным фактором ростовых процессов является свет , под влиянием которого уменьшается рост стеблевых органов, увеличивается их дифференциация, вследствие этого происходит утолщение стебля. При недостатке света, и особенно при полном затемнении, наблюдается сильное удлинение стебля, увеличивается длина междоузлий и уменьшается их толщина.
- Большое влияние на рост побега оказывает приток кислорода воздуха, который необходим для дыхания быстро делящихся клеток верхушечной меристемы в конусе нарастания, а также усиления деятельности камбия.
При недостатке кислорода рост побега замедляется.
- На рост побегов оказывает влияние и ряд других факторов.
Как известно, близость побега к корневой системе обуславливает большую энергию роста. В связи с этим побеги, выросшие из сучка замещения или из многолетней части, всегда растут интенсивнее, чем побеги на плодовой стрелке (К. Д. Стоев и Б. Рангелов, 1969). Побеги, развившиеся из различных почек по длине стрелки, также растут неравномерно. Как правило, рост проходит интенсивнее у побегов находящихся у основания и в конце плодовой стрелки. Так, для сорта Мискет червен установлены 2 максимума роста побегов - первый в секторе 3-4-го глазка, второй - в секторе 10-12-го глазка плодовой стрелки.
- Между ростом отдельных побегов, их частей и других органов существует определенная взаимосвязь и взаимозависимость. Их изучение имеет большое значение для обоснования агротехнических приемов, применяемых с целью регулирования роста и плодоношения, как например, обрезка, зеленые операции, подвязка и т.п. В процессе роста отмечена связь между верхушкой побега и его боковыми точками роста. Растущая верхушка побега тормозит пробуждение боковых точек роста, что можно объяснить тормозящим влиянием ростовых стимуляторов - ауксинов, вырабатываемых растущим побегом. При удалении верхушки основного побега трогаются в рост пасынки.
Существует зависимость между каждым растущим побегом и всеми остальными побегами на кусте. Если сильно растущие побеги прищипнуть, и, таким образом, остановить их рост, другие побеги растут интенсивнее.
Между ростом надземной и подземной частями куста наблюдается связь, что имеет значение для теоретического обоснования обрезки. Так, ежегодная, короткая обрезка сокращает листовую поверхность и уменьшает накопление пластических веществ. Однако, корневая система требует для своего развития все большего количества питательных веществ, а когда этих веществ будет недостаточно для увеличения числа корней, часть молодых корешков отомрет. Угнетение корневой системы, в свою очередь отражается на развитии надземной части, что приводит к еще большему угнетению роста побегов. Ослабление роста корневой системы сильнее влияет на рост надземной части куста, чем ослабление роста надземной части на рост корневой системы, что объясняется различием в их питании.
Повреждение корневой системы немедленно сказывается на росте надземной части, тогда как корни оставленные без надземной части, могут оставаться живыми в течение нескольких лет, так как в них накапливается в больших количествах запасные вещества.
Наблюдается прямая связь между силой роста корневой системы и числом побегов на кусте. Установлено, что чем больше развита корневая система, тем больше образуется побегов.
- Транспирация и ассимиляция побегов. Различают транспирацию устьичную и кутикулярную. У винограда транспирация листьев осуществляется главным образом через устьица. На зеленом побеге устьиц значительно меньше, чем на листе, поэтому транспирация побегов слабая и главным образом, кутикулярная. Верхушечные травянистые части побегов транспирируют сильнее, чем более старые (средние или нижние), и поэтому при срезании, увядают сначала самые верхушечные молодые части побега. Однолетние вызревшие побеги испаряют воду значительно слабее, так как они покрыты слоем пробковой ткани.
Ассимиляция углерода у побегов слабая, так как они содержат хлорофилл в значительно меньшем количестве, чем листья, а также из-за небольшого числа устьиц в них и более толстого эпидермиса.
Химический состав побега изменяется в зависимости от сорта, фазы вегетации, климатических и почвенных условий произрастания.
Главными химическими составными элементами являются вода, органические и минеральные соединения.
Содержание воды в стебле несколько меньше, чем в корне. В одревесневших побегах содержание воды колеблется от 30 до 52 %, а в зеленых - от 55 до 75%.
Органические вещества составляют основную массу соединений, входящих в состав протопласта и оболочки. В оболочках клеток содержится целлюлоза, гемицеллюлоза и пентозаны. В протопласте клеток имеются разнообразные органические вещества, многие из которых служат запасными, например крахмал, сахар, жиры, дубильные вещества, щавелевокислые соли и др.
В свежих зеленых побегах винограда у разных сортов V. vinifera содержится 0,90-1,02% сырого жира; 6,85-15,66% безазотистых экстрактивных веществ.
Сырой белок в зеленых побегах с листьями составляет 3,49-4,05%. В конце вегетации (по данным Я. И. Потапенко и Е. И. Захаровой), в однолетних побегах винограда содержалось моносахаров 7,23%, дисахаров - 5,14% и крахмала 8,00% на абсолютно сухое вещество.
Содержание минеральных веществ в органах винограда устанавливается на основании анализа золы. Вызревшие побеги содержат от 2,3 до 3,7 г золы на 100 г абсолютно сухого вещества. В золе больше всего кальция (1/3-1/2 часть всего состава золы), второе место занимает калий (1/4-1/3 часть всей золы), меньше содержится фосфора (примерно 1/10 часть).
Молодые части побега богаче калием и фосфором, а в старых побегах преобладает кальций и магний.
Хлорофилл в значительном количестве содержится в одно- и двухлетних побегах, а в четырехлетних и старше частях куста хлорофилл имеется только в коре и в молодых слоях древесины.
Установлено, что в побегах виноградной лозы имеется витамин С, максимальное содержание которого отмечено перед созреванием винограда.
Рост и вызревание побегов виноградной лозы сопровождается активным обменом углеводов, белков, дубильных веществ, органических кислот и т.д.
Исследованиями К. Д. Стоева (1948) установлено, что в содержании сахаров и крахмала в однолетних побегах имеются два максимума и два минимума. Первый максимум содержания крахмала отмечается в конце марта - начале апреля, когда передвинувшиеся из корневой системы к почкам винограда сахара временно откладываются в форме крахмала. Этому периоду соответствует сильное понижение содержания сахаров в побеге, что, по-видимому, объясняется их ресинтезом в крахмал, после того как будут исчерпаны их защитные функции во время зимовки винограда. В период интенсивного роста побегов происходит непрерывное уменьшение содержания крахмала и повышение содержания сахаров, максимум которых отмечается в разгар цветения.
Второй максимум содержания крахмала обнаруживается в конце октября - начале ноября, т.е. в конце вегетационного периода, в фазу вызревания побегов. К этому времени (а в средних и нижних зонах побега несколько раньше) отмечен минимум в содержании сахаров. После листопада наступает интенсивный гидролиз крахмала, и, одновременно с ним, сильное повышение содержания сахаров, что вызвано коренным изменением функции пластических углеводов в жизни винограда.
В своих исследованиях И. Н. Кондо (1959) установил, что с возрастом побегов обогащенность запасами подвижных углеводов резко уменьшается. Однолетний побег сорта Саперави содержит 16,87% углеводов (сахар + крахмал), что вдвое больше, чем восьмилетние побеги (7,6%).
- Содержание белковых и азотистых веществ в побегах винограда претерпевает некоторые изменения в течении периода вегетации.
Исследованиями Е. А. Макаревской (1966) установлено, что во второй половине лета и в начале осени, параллельно уменьшению содержания редуцирующего сахара в виноградных побегах наступает повышение содержания белкового и общего азота в коре и уменьшается количество растворимого небелкового азота. Зимой, при выходе почек из состояние покоя, отмечается новое повышение белкового азота. У более зимостойких сортов содержание азотистых веществ несколько выше. Исследованиями К. Д. Стоева и др. (1960) в побегах виноградной лозы во время их вызревания и покоя установлены следующие свободные аминокислоты и амиды: гистидин, аспарагин, аспарагиновая кислота, глютаминовая кислота, треонин, аланин, тирозин, норвалин, изолейцин, пролин, валин.
Б. Л. Африкян и др. (1962) установил, что в результате естественного или искусственного промораживания побегов у неморозоустойчивого сорта, аминокислотный состав белков подвергается значительным изменениям. Это наблюдается, в незначительной степени, и у морозостойкого сорта. Весной аминокислотный набор морозостойкого сорта остается почти без изменений, тогда как у неморозостойкого сорта их количественный и качественный состав значительно меняется. Это дает основание автору полагать, что аминокислотный состав белков имеет определенное отношение к морозоустойчивости сортов.