Начало цветения отмечается по опадению первых т. наз. колпачков венчика. Кусты различных сортов, различные соцветия одного куста и различные цветки одного и того же соцветия цветут не одновременно. Раньше всего цветут, как правило, североамериканские виды винограда и сорта подвойные и прямые производители. Европейско-азиатские сорта, как правило, цветут позднее. Ранние из них по распусканию почек и созреванию (например, Жемчуг Саба, Фетяска и др.) цветут раньше поздних сортов (Гымза, Фурминт). Позднее всего, как правило, цветут крупноягодные столовые и изюмные сорта восточного происхождения (Паркент, Карабурну, Султанина и др.).
Нормальная последовательность цветения сортовых групп и сортов получается только в случае раннего наступления весны и благоприятной весенней погоды. Поздняя весна и задержка распускания почек, внезапное потепление нарушают последовательность цветения сортов, ввиду чего ранние и поздние сорта цветут почти одновременно.
Начало цветения сорта связано со сроком распускания почек. Кусты, почки которых распускаются раньше, зацветают раньше. По некоторым наблюдениям отдельные сорта ежегодно цветут после образования определенного числа междоузлий (Мержаниан, 1951; Коваль-Мартьянова, 1963). Например, Витис рипария, Рипария х Рупестрис 101-14, Бако № 1, Зейбель. № 880, 4995, 4986 зацветают после образования 7—9-го междоузлий; Изабелла, Ноа, Отелло, Лидия — 9—11-го междоузлий; Чауш, Оппорто, Ранний Маленгра, Жемчуг Саба — 13—14-го междоузлий; Шасла белая, Алиготе, Мускат гамбургский, Красавица Цегледа, Алимшак, Сенсо, Каберне Совиньон, Фетяска, Коарнэ нягрэ—14—16-го междоузлий; Карабурну, Италия, Агадаи — 15—17-го, а сорт Тавриз — после образования 18-20-го междоузлий.
Соцветия на побегах, расположенных выше, начинают цвести раньше, чем на более низких побегах. Соцветия на более низкой части побега также цветут раньше, чем расположенные выше ( anаrеsi, 1957; Мержаниан, 1951). На кустах с низким штамбом и на солнечных склонах цветение начинается раньше, чем на высокоштамбовых кустах и на влажных, холодных почвах. Цветение более крупных соцветий наступает раньше, чем соцветий с меньшим числом цветков.
В условиях благоприятной погоды цветение одного соцветия протекает за 4-8 дней, сорта — за 8—14, а насаждения — за 12—20 дней.
Фазу цветения можно разделить на три периода: а) начало цветения; б) разгар цветения; в) конец цветения. Началом цветения считается момент раскрывания 4—5 % цветков. В разгар цветения цветет 60—70 % цветков. В конечный период цветения цветет 25—30% цветков. Начальный и конечный периоды цветения более растянуты.
Важнейшим экологическим фактором начала и протекания цветения является температура; кроме того, важную роль играют осадки и влажность воздуха.
Для начала цветения различные по сроку созревания сорта требуют различных сумм активных температур. По данным Мержаниана (1951), от распускания почек до начала цветения требуется сумма активных температур в 380°. По Иванову (1952) Жемчуг Саба начинает цвести после действия 576° эффективных и 190° активных, Ранний, Маленгра — 607° эффективных и 185° активных, Шасла белая — 632° эффективных и 207° активных, Алиготе — 594° эффективных и 191° активных, Papa нягра — 629° эффективных и 207° активных температур.
По более новым исчислениям Homey (1966), для начала цветения Рислинг райнский требует 8000, Сильванер 7900, Мюллер-Тургау 7600 градусо-часов.
Экологические факторы цветения были изучены уже многими исследователями, которые установили, что сорта винограда начинают цвести при температуре 16—25 °С, а оптимальной для цветения является температура в 20— 26°С (Viala-Pechoutre, 1910; Sartorius, 1926; Мержаниан, 1951 и др.). По наблюдениям в Венгрии (Коzmа, 1951, 1951а) при температуре 10—13°С раскрываются только отдельные цветки, а при 13—17°С цветение не спонтанное.
Слишком высокая температура, сопровождаемая низкой относительной влажностью воздуха, также мешает нормальному ходу цветения. По наблюдениям Constantinescu с сотр. (1952), температура 30—40°С во время цветения вызывает массовое высыхание и бесплодие цветков, особенно если относительная влажность воздуха ниже 45%. Относительная влажность воздуха ниже 40% даже при оптимальной температуре подавляет цветение. Для цветения нежелательна и влажная погода с осадками. При такой погоде даже в случае оптимальной температуры раскрывание бутонов не будет спонтанным, поскольку отчленение колпачков идет медленно и большая часть их не отделяется от пестика даже после отрыва их от основания.
Цветению винограда свойственна своеобразная суточная эндогенная периодичность (Sartorius, 1926; Мержаниан, 1951; Constantinescu с сотр., 1952; Коша, 1951, 1951а). Распускание бутонов начинается в 6-8 ч. утра, и количество раскрывшихся бутонов возрастает до 9-10 ч., после чего к 10-11 ч. интенсивность раскрывания затухает и наконец приостанавливается. Во второй половине дня, около 15—16 ч., вновь наступает короткий период распускания бутонов. Ночью бутоны не открываются (рис. 1). Индуцированная периодичность цветения была открыта Sartorius (1926) и подтверждена Коzmа (1951, 1951а). Это явление заключается в том, что на интенсивность раскрывания бутонов влияют температурные условия предыдущего дня. Если погода предыдущего дня была ясная и теплая, то интенсивность раскрывания бутонов даже при небольшом снижении температуры будет более высокая, чем при прохладной погоде предыдущего дня и потеплении в день наблюдения (рис. 2). Изменения света и влажности воздуха на периодичность цветения особого влияния не имеют.
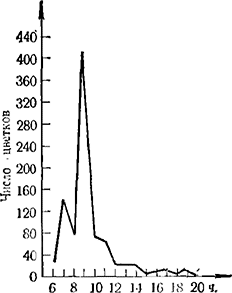
Рис. 1. Суточная периодичность цветения
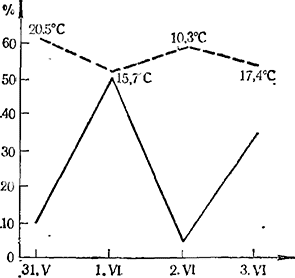
Рис. 2. Интенсивность цветения (%) в зависимости от температуры
Наблюдениями Мержаниана (1951) установлено, что чем позже начинается цветение соцветия, тем быстрее оно заканчивается. Во второй половине цветения суточный максимум раскрывания бутонов наступает в более ранние утренние часы.
Интенсивнее всего цветение идет при температуре 25—35°С. Чем интенсивнее утреннее потепление и быстрее снижение относительной влажности воздуха, тем быстрее проходит цветение и короче его суточный период. Чем ниже средняя температура дня и выше относительная влажность воздуха и чем она меньше изменяется в течение дня, тем более растянутым будет период цветения (Коzmа, 1951). Kozma показал также, что бутоны подготовляются к распусканию в предыдущий день, на что им требуется 7—12 h.
Распускание цветков происходит следующим образом. По окончании роста цветков и образования пыльцы и зародышевого мешка за несколько дней до цветения у основания лепестков, в месте соприкосновения их с диском, образуется разделяющий слой ткани (Sartorius, 1926; Мержаниан, 1951), который состоит из неплотно расположенных, округлых и удлиненных клеток. Разделяющий слой сначала образуется в одном лепестке, а потом образование его распространяется на соседние лепестки. Иногда можно различить первичные и вторичные разделяющие слои клеток (Мержаниан, 1951). В этих слоях накопляется большое количество крахмала. Одновременно с образованием разделяющего слоя ткани лепестки приобретают желтовато-зеленый цвет и постепенно прекращается поступление к ним водорастворимых питательных веществ.
В день цветения под воздействием повышения температуры и снижения влажности воздуха в наружных клетках лепестков влажность снижается быстрее, нежели во внутренних, и поскольку приток влаги к ним уже прекращен, возникает напряжение, в результате чего от основания отрывается сначала один лепесток венчика, потом через 20—30 s подряд остальные лепестки и, наконец, венчик отчленяется в виде колпачка. Отчленению венчика способствуют набухшие диски — нижний и верхний диски или нектарник (Баранов, 1946). После этого тычиночные нити быстро удлиняются и отходят от пестика под углом 45°, отчего колпачок отрывается от цветка. После отпадения венчика тычиночные нити принимают характерное для типа цветка положение. Пыльники открываются в процессе сбрасывания колпачка или после него. При растянутом цветении (в случае прохладной, влажной погоды) пыльники открываются в процессе сбрасывания колпачка или перед сбрасыванием, а при быстром цветении (в случае теплой, сухой погоды) они открываются и выбрасывают пыльцу спустя 1—60 min после сбрасывания колпачка. Раскрывание пыльника вызывается также быстрой отдачей воды его оболочкой.
Тычиночная нить остается живой приблизительно день после раскрывания цветка, буреет, подсыхает и наконец отмирает. Через некоторое время после раскрывания бутонов на рыльце появляется капелька жидкости величиной с булавочную головку, называемая секреторной жидкостью, функция которой заключается в создании среды для прорастания пыльцевых зерен.
Появление секреторных капель, так же как цветение, имеет своеобразный суточный эндогенный ритм. Как правило, капли массово появляются в ранние утренние часы, кроме того, они могут иметь еще один период появления, во второй половине дня (в 17—18 ч.). Секреторные капли ежедневно ритмично появляются до тех пор, пока цветок не оплодотворится или же при отсутствии оплодотворения рыльце не побуреет. Рыльце стареет через 10—12 дней после распускания цветков, что видно и по его побурению и подсыханию. После прекращения появления секреторных каплей у основания цветоложа образуется разделяющий слой клеток и начинается осыпание цветков (Kozma, 1951).
При тепличной культуре винограда на цветках появляется такое количество секреторных капель, что они стекают с рыльца и вместе с ними стекают и пыльцевые зерна, попавшие на рыльце.
Важная роль секреторных капель в развитии пыльцевых трубок в процессе оплодотворения подчеркнута несколькими исследователями. При сухой погоде секреторная жидкость высыхает; пыльца не в состоянии развивать трубку (Мержаниан, 1919, 1929, 1951). Branscheidt (1929) различает секреты рыльца от экскреций пыльцевой трубки и доказывает, что по химическому составу они не одинаковы. Оба секрета присутствуют в секреторной жидкости рыльца, в случае высыхания она непригодна для развития пыльцевой трубки. Секреторные капли выделяют лишь обоеполые и функционально женские цветки, концентрация их в зависимости от внешних и внутренних условий может быть различной, а их отсутствие может вызвать осыпание (Rathау, 1888). Секреторная деятельность функционально женского цветка более интенсивна, нежели обоеполого цветка (Ziegler, Branscheidt, 1927). На рыльце цветков мужского характера, звездчатых и других аномальных цветков секреторной жидкости не выделяется (Kozma, 1961, 1961а,b,с, 1963).
В оплодотворении цветка винограда большую роль играют биохимическое состояние цветка и обмен веществ цветка.
Раздельнополые цветки дикого винограда ( V. vinifera silvestris Gmеl)*, как правило, а обоеполые цветки большинства сортов культурного винограда *
(V. vinifera L.) большей частью цветут при полном сбрасывании венчика (хазмогамия). Существуют сорта, у которых при цветении венчик не сбрасывается, а остается на пестике (клейстогамия). В этом случае оплодотворение происходит исключительно автогамно. Клейстогамно могут оплодотворяться только сорта с фертильной пыльцой, а сорта с функционально женским типом цветка таким путем могут образовать лишь бессемянные ягоды. Некоторые обоеполые сорта (например, Медок черный, Эзеревеш Мадярорсаг эмлеке, Мускат Матяш янош) склонны к клейстогамии, и большинство их цветков оплодотворяется таким путем. Клейстогамное оплодотворение может происходить и в результате заражения патогенными грибками, солнечного ожога или ожога ядохимикатами. После цветения в ходе роста ягод лепестки разрушаются, высыхают и опадают.
* Проф. Р. Kozma придерживается точки зрения, согласно которой европейско-азиатский виноград относится к V. vinifera L., а дикий виноград, распространенный в лесах, — к silvestris Gnаеl.